Miyacha
Miyacha (lot. cerebellum – kichik miya) umurtqalilar bosh miyasining bir qismi. Odatda miya yarimsharlaridan kichik, lekin ba’zi baliqlarda undan katta boʻlishi ham mumkin.[1] Odamlarda miyacha harakat boshqariluvida katta rol oʻynaydi. U shuningdek ba’zi bir kognitiv funksiyalarda, masalan diqqat va til, shuningdek qoʻrquv va qoniqish kabi hissiy boshqaruvlarda ham ishtirok etadi,[2][3] ammo asosiy ta’siri harakatga bogʻliq funksiyalarda. Odam miyachasi harakatni chaqirmaydi, lekin harakatdagi koordinatsiya, tartib va aniq davomiylikni ta’minlaydi: u orqa miya va miyaning boshqa qismlaridan sezuvchi impulslarni qabul qiladi va ular asosida harakatning oʻta aniq chiqishini ta’minlaydi.[4] Miyachaning jarohatlanishi odamlarda nozik harakatlar, tana holati boshqariluvi va yangi harakatlar oʻrganilishi buzilishiga olib keladi.[4]
Anatomik jihatdan, odam miyachasi bosh miyaning pastki qismi tegib turuvchi alohida tuzilma boʻlib, miya katta yarimsharlari pastida yopishib turadi. Uning poʻstlogʻi tartibsiz burmalarga ega yarimsharlar poʻstlogʻidan farq qilib, parallel egatlardan iborat. Bu parallel egatlar miyacha poʻstlogʻi aslida akkordion holatiga keltirilgan davomli yupqa toʻqima ekanligini koʻrsatadi. Bu yupqa qatlamda bir qancha neyron turlari oʻzaro tartib bilan joylashgan. Eng muhimlari Purkinye hujayralari va donador hujayralarder. Bu kompleks tuzilma katta miqdordagi signallar bilan ishlash imkoniyatini beradi, lekin miyacha poʻstlogʻidan chiquvchi deyarli barcha ma’lumot miyacha ichidagi oq moddada joylashgan kichik chuqur yadrolar tizimi orqali oʻtadi.[5]
Miyacha harkat boshqaruvidagi asosiy rolidan tashqari, harakatni oʻrganishning bir qancha turlarida muhim, sezgi-harakat (sensorimotor) munosabatlari oʻzgarishlarida yangi harakat oʻrganishdagi ishtiroki sezilarli. Sezgi-harakat kalibratsiyasini miyachadagi [sinaptik plastiklik]ka bogʻliqligini koʻrsatuvchi bir qancha nazariy modellar ishlab chiqilgan. David Marr va James Albus tomonidan ilgari surilgan modellar har bir miyacha Purkinye hujayralari umuman farqli 2 xil signalni qabul qilishini oʻrganish asosida yaratilgan: biri donador hujayralarning parallel tolalaridan keluvchi minglab kuchsiz signallar; boshqasi yagona koʻtariluvchi toladan keluvchi juda kuchli signallarni qabul qilish.[6] Marr–Albus nazariyasining asosiy ma’nosi koʻtariluvchi tolalar “oʻrgatuvchi signal”larni tashiydi va parallel tolalardagi signal kuchini uzoq davom etuvchi oʻzgarishini chaqiradi. Parallel tolalardagi uzoq muddat depressiyalarini oʻrganish shu turdagi nazariyalarni rivojlantirdi, lekin ularning ishonchliligi baxsli masala.[7]
Tuzilishi
[tahrir | manbasini tahrirlash]Umumiy anatomiyaga koʻra, miyacha zich poʻstloq, uning ostidagi oq modda va asosidagi suyuqlik bilan toʻlgan qorinchadan iborat. Toʻrtta miyacha chuqur yadrolari oq modda ichida joylashgan. Poʻstloqning har bir qismi yuqori geometrik stereotip bilan joylashgan kichik neyronal element tuzilmalaridan iborat. Miyacha va uning yordamchi tuzilmalari bir necha yuzlab yoki minglab mustaqil ishlovchi, “mikrozona” yoki “mikroqismlar” deb ataluvchi modullarga boʻlinishi mumkin.
Umumiy anatomiya
[tahrir | manbasini tahrirlash]
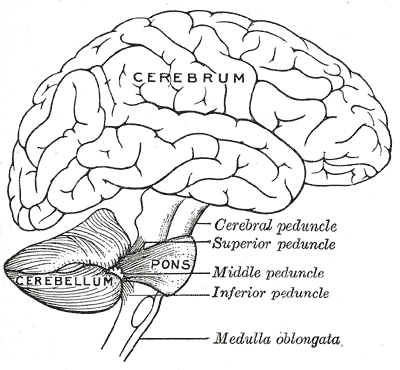
Miyacha kalla suyagining orqa chuqurchasi da joylashgan. Toʻrtinchi qorincha, miya koʻprigi va uzunchoq miya miyachaning oldingi tomonida joylashgan.[8] U yuqorida yotuvchi bosh miyadan dagʻal qattiq parda bilan ajralib turadi; Uning miyaning boshqa qismlari bilan qolgan barcha aloqalari koʻprik orqali oʻtadi. Anatomlar miyachani metencephalonning bir qismi deb tasniflaydilar, unga koʻprik ham kiradi; metencephalon rombencephalonning yuqori qismi hisoblanadi. Bosh miya poʻstlogʻiga oʻxshab, miyacha ham ikki miyacha yarimsharlariga boʻlinadi; unda yana tor oʻrta zona chuvalchangsimon qism (vermis) ham bor. Katta burmalar tizimi, kelishuvga koʻra umumiy strukturani 10 ta kichikroq “boʻlakchalar”ga boʻladi. Undagi koʻplab mitti donador hujayralar hisobiga miyacha miyaning qolgan barcha qismlari umumlashtirilganidan koʻra koʻproq neyronlar tutadi, lekin u butun miya hajmining 10% inigina egallaydi.[9] Miyachadagi neyronlar soni neokorteksdagi neyronlar soniga bogʻliq. Miyachada neokorteksdagidan 3,6 marta koʻp neyronlar bor va bu nisbat turli xil sutemizuvchilarda bir xilligi kuzatilgan.[10]
Miyachaning noodatiy yuzasi uning asosiy hajmi zich taxlangan kulrang modda qavati: miyacha poʻstlogʻidan iboratligini yashirib turadi. Bu qavatdagi har bir pushta folium deb ataladi. Hisoblanishicha, odam miyachasi poʻstlogʻi toʻliq yozib tekislansa, 1 metr uzunlik va 5 santimetr kenglikda, umumiy 500 sm kvadrat maydonni egallaydi, hajmiy birliklarga qoʻysak 6 sm × 5 sm × 10 sm boʻladi.[9] Poʻstloqning kulrang moddasi ostida oq modda yotadi. U poʻstloqqa boruvchi va undan ketuvchi koʻplab miyelinlangan nerv tolalaridan iborat. Oq modda ichida toʻrtta miyacha chuqur yadrolari joylashgan. Ular koʻndalang kesmada daraxt shoxlarini eslatgani uchun arbor vitae (hayot daraxti) deb ataladi.[11]
Miyacha nerv sistemasining turli qismlari bilan miyacha oyoqchalari (lot.pedunculi cerebellaris) orqali bogʻlangan. Bular yuqori (pedunculi cerebellaris superior), oʻrta (pedunculi cerebellaris medius) va pastki (pedunculi cerebellaris inferior) miyacha oyoqchalari boʻlib, chuvalchangsimon qismga nisbatan joylashuvi boʻyicha nomlangan. Yuqorigi miyacha oyoqchalari thalamus yadrolari orqali yarimsharlar poʻstlogʻidagi yuqorigi motor neyronlarga boruvchi efferent tolalar tutadi. Tolalar miyacha chuqur yadrolaridan chiqadi. Oʻrta miyacha oyoqchalari koʻprik bilan bogʻlangan va barcha impulslarni koʻprikdan, asosan nucleus pontineusdan oladi. Yarimsharlar poʻstlogʻidan koʻprikka oʻtgan impulslar ushbu yadro va uning tolalari orqali miyachaga oʻtkaziladi. Oʻrta oyoqcha oyoqchalar ichida eng yirigi boʻlib, miyachaning turli qismlariga boruvchi uchta tutamlarga ajratiladi. Pastki miyacha oyoqchalari vestibulyar yadrolar, orqa miya va koʻprik tomi (lot. tegmentum) dan impulslar qabul qiladi. Pastki oyoqchadan chiquvchi impulslar efferent tolalar orqali vestibulyar yadrolar va retikulyar formatsiyaga boradi. Butun miyacha boshqaruvchi impulslarni pastki miyacha oyoqchalari orqali pastki oliva yadrosi (lot. nucleus olivaris inferior)dan oladi.[5]
Boʻlaklari
[tahrir | manbasini tahrirlash]
Yuzasiga koʻra miyacha uch boʻlakka boʻlinishi mumkin: miyachaning oldingi (lot. lobus cerebelli anterior) (birlamchi egat ustida), orqa (lot. lobus cerebelli posterior) (birlamchi egat ostida) va boʻlakcha-tuguncha (lot. lobus flocculonodularis) boʻlaklariga. Bu boʻlaklar miyachani rostral va kaudal (odamlarda yuqorigi va pastki) qismlarga boʻladi. Funksiyasi jihatidan medialdan-lateralga shaklida boʻlish muhim. Flocculonodular boʻlakdan tashqari (uning oʻz bogʻlanishlari va funksiyalari bor) miyachani funksional jihatdan spinocerebellum deb ataluvchi medial sektor va cerebrocerebellum deb ataluvchi lateral sektorga boʻlish mumkin.[11] Oʻrta chiziqdan chiqib turuvchi uzun va tor boʻlak miyachaning chuvalchangsimon qismi deb ataladi.[11]
Eng kichik soha, flocculonodular boʻlak, odatda vestibulocerebellum deb ataladi. U evolyutsion jihatdan eng qari (archicerebellum) qism va asosan muvozanat va makon oriyentatsiyasida ishtirok etadi; uning asosiy aloqalari vestibulyar yadrolar bilan, shuningdek vizual va boshqa sezgi impulslarini ham qabul qiladi. Bu sohaning zararlanishi qadam tashlash va muvozanat buzilishiga sabab boʻladi.[11]
Oldingi va orqa boʻlaklarning medial zonalari spinocerebellum, shuningdek paeocerebellum deb ham ataladi, tashkil qiladi. Miyachaning bu sektori tananing va qoʻl-oyoqlarning nozik harakatlarida ishtirok etadi. U orqa miyaning orqa ustunlaridan spinocerebellar tract bilan birga proprioseptiv impulslarni va bosh miya nervlaridan uch shoxli nerv, shuningdek vizual va eshitish sistemalaridan ham impulslar qabul qiladi.[12] U miyacha chuqur yadrolariga tolalar joʻnatadi, oʻz navbatida, yarimsharlar poʻstlogʻi va miya ustuniga proyeksiyalanishi orqali tushuvchi harakat sistemalari modulyatsiyasini ta’minlaydi.[11]
Lateral zona odamlarda eng katta qism boʻlib, cerebrocerebellumni tashkil qiladi, shuningdek neocerebellum deb ham ataladi. U yarimsharlar poʻstlogʻidan, xususan chakka boʻlakdan nucleus pontineus orqali (cortico-ponto-cerebellar yoʻllarni hosil qilib) impulslarni qabul qiladi va oldingi lateral talamusga (oʻz navbatida yarimsharlar poʻstlogʻining premotor va birlamchi harakat sohasiga) va qizil yadroga impuls yuboradi.[11] Lateral miyachaning funksiyalarini ta’riflashda kelishmovchiliklar mavjud: u bajariladigan harakatni rejalashtirishda,[13] harakat uchun sensor axborotlarni baholashda,[11] va bir qancha kognitiv funksiyalarda, masalan qaysi fe’l ma’lum bir otga eng mos kelishi (masalan, "oʻtirish" "stul" uchun)ni aniqlashda ishtirok etadi deb hisoblanadi.[14][15][16][17]
Mikroanatomiya
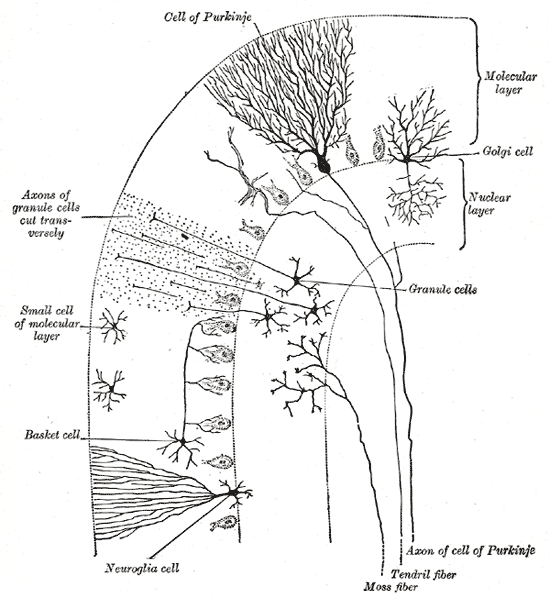
[tahrir | manbasini tahrirlash]Miyachada ikki neyron turi asosiy rol oʻynaydi: Purkinye hujayralari va donador hujayralar. Uch turdagi aksonlar ham muhim rol oʻynaydi: moxsimon tolalar, koʻtariluvchi tolalar (miyachaga tashqaridan kiradi) va parallel tolalar (donador hujayralarning aksonlari). Miyacha ichida ikkita asosiy yoʻl boʻlib, moxsimon va koʻtariluvchi tolalardan hosil boʻlgan va ikkovi ham miyacha chuqur yadrolariga boradi.[9]
Moxsimon tolalar toʻgʻridan-toʻgʻri chuqur yadrolarga boradi, lekin quyidagi yoʻnalish orqali ham yuradi: moxsimon tolalar → donador hujayralar → parallel tolalar → Purkinye hujayralari → chuqur yadrolar. Koʻtariluvchi tolalar Purkinye hujayralariga boradi, shuningdek toʻgʻridan-toʻgʻri chuqur yadrolarga ham kollaterallar beradi.[9] Moxsimon va koʻtariluvchi tolalar har biri maxsus axborotlarni tashiydi; miyacha shuningdek dofaminergik, noradrenergik, serotoninergik va xolinergik impulslarni ham qabul qiladi va global modulatsiyani ta’minlaydi.[18]
Miyacha poʻstlogʻI uch qavatga boʻlingan. Eng pastda qalin donador qavat, zich joylashgan donador hujayralar, shuningdek interneyronlar, asosan Golji hujayralari shuningdek Lugaro hujayralari va unipolyar choʻtkasimon hujayralardan tashkil topgan. Oʻrtada Purkinye qavati yotadi. U Purkinye hujayralari va Bergmann glial hujayralaridan iborat ingichka qavat. Eng yuqorida molekulyar qavat yotadi va Purkinye hujayralarining tekislangan dendritik daraxtlari, shuningdek Purkinye hujayralari dendritik daraxtlariga toʻgʻri burchak ostida kirgan bir qator yirik parallel tolalardan tashkil topgan. Miyachaning bu eng tashqi qavatida shuningdek ikki turdagi ingibitsiyalovchi interneyronlar boʻladi: yulduzsimon hujayralar va savatsimon hujayralar. Ikkala hujayralar ham Purkinye hujayralar dendritlari bilan GAMKergik sinapslar hosil qiladi.[9]

Andoza:Bull(-): Ingibirlovchi birikish
Andoza:BullMF: moxsimon tolalar
Andoza:BullDCN: Miyacha chuqur yadrolari
Andoza:BullIO: Pastki oliva yadrosi
Andoza:BullCF: Koʻtariluvchi tola
Andoza:BullCFC: Koʻtariluvchi tola kollaterali
Andoza:BullGC: Donador hujayralar
Andoza:BullPF: Parallel tola
Andoza:BullPC: Purkinye hujayrasi
Andoza:BullGgC: Golji hujayrasi
Andoza:BullSC: Yulduzsimon hujayra
Andoza:BullBC: Savatsimon hujayra
Purkinye hujayralari
[tahrir | manbasini tahrirlash]
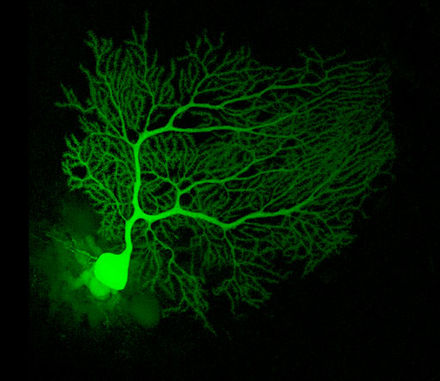
Purkinye hujayralari miyadagi eng farqlanuvchi neyronlardan boʻlib, eng erta aniqlanganlaridandir – ular birinchi marta chex anatomi Jan Evangelista Purkyně tomonidan 1837-yilda tasvirlab berilgan. Ular oʻzlarining dendritic daraxtlari shakliga koʻra aniqlanadi: dendtritlar juda kuchli tarmoqlangan, lekin miyacha pushtalariga nisbatan qat’iy perpendikulyar tekislanib joylashgan. Shunday qilib Purkinye hujayralari dendritlari zich planar tarmoq hosil qiladi va ular orasidan parallel tolalar toʻgʻri burchak ostida oʻtadi.[9] Dendritlar dendrit oʻsiqchalari bilan qoplangan va ularning har biri parallel tolalardan sinaptik impuls qabul qiladi. Purkinye hujayralari miyadagi boshqa har qanday turdagi hujayradan koʻra koʻproq sinaptik boglanishga ega – odamning bitta Purkinye hujayrasida taxminan 200 000 atrofida oʻsiqchalar boʻladi.[9] Purkinye hujayralarining katta, sferik tanalari miyacha poʻstlogʻidagi bir hujayra qalinligidagi tor qavatga joylashtirilgan va “Purkinye qavati” deb ataladi. Ularning aksonlari poʻstloqning yaqin sohalariga kollaterallar bergach, miyacha chuqur yadrolariga boradi va bu yerda bir qancha turdagi yadro hujayralari bilan 1000 atrofida bogʻlanishlar (hammasi bilan kichik domenlarda) hosil qiladi. Purkinye hujayralari neyromediator sifatida GAMKni ishlatadi va shuning uchun nishonlariga ingibirlovchi ta’sir koʻrsatadi.[9]
Purkinye hujayralari miyacha yoʻllarining yuragini tashkil qiladi. Ular oʻlchamining kattaligi va oʻziga xos faoliyati tufayli hayvonlarda ekstrasellulyar yozib olib usullari bilan oʻrganishda qulayliklar yaratadi. Purkinye hujayralari normada hech qanday sinaptik impuls kelmasa ham katta chastotada harakat potensiallari tarqatib turadi. Uygʻoq hayvonlarda taxminan 40 Hz atrofida. Harakat potensiallarining chizmalari oddiy va murakkab spayk (ingl. spike)lar aralashmasidan iboratligini koʻrsatadi. Oddiy spayk bitta harakat potensiali va undan keyin kelgan taxminan 10 ms davom etuvchi refraktorlik davridan iborat; murakkab spayk juda qisqa spayklararo intervallar va pasayib boruvchi amplitudali harakat potensiallarining bir xil zanjiridan iborat.[20] Fiziologik oʻrganishlar shuni koʻrsatadiki, murakkab spayklar (bazal (ing. baseline) chastotalari 1 Hz atrofida va hech qachon 10 Hzdan oshmaydi) koʻtariluvchi tolalar aktivlashishiga bogʻliq. Oddiy spayklar boʻlsa bazal aktivlik va parallel tollalardan keluvchi impulslar kombinatsiyasidan hosil boʻladi. Murakkab spayklar odatda oddiy spaklar faoliyati susaygan bir necha yuz millisekundlarda keladi.[21]
Purkinye neyronlarining maxsus, oʻziga xos xususiyati kalbindin ekspressiyasidir.[22] bir tomonlama oʻtirgʻich nervining surunkali jarohati boʻlgan kalamush miyasini kalbindinli boʻyash shuni koʻratdiki, voyaga yetgan miyada Purkinye hujayralari yangidan hosil boʻlishi va miyachada yangi boʻlakchalar hosil boʻlishiga olib kelishi mumkin.[23]
Donador hujayralar
[tahrir | manbasini tahrirlash]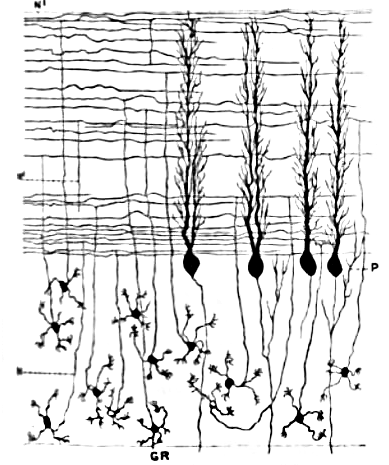
Miyacha donador hujayralari, Purkinye hujayralaridan farqli miyadagi eng kichik neyronlarga kiradi. Ular shuningdek miyadagi eng soni koʻp neyronlardir: odamlarda umumiy soni taxminan 50 mlrd, bu miyadagi neyronlarning ¾ qismi miyacha donador hujayralari degani.[9] Ularning hujayra tanalari miyacha poʻstlogʻi pastki qismidagi qalin qavatda joylashgan. Donador hujayra toʻrt yoki beshtagina dendritga ega, ularning har biri “dendrit changali” deb ataluvchi kengaymada tugaydi.[9] Bu kengaymalar moxsimon tolalardan keluvchi qoʻzgʻatuvchi impulslar va Golji hujayralaridan keluvchi ingibirlovchi impulslarni qabul qiladi.[9]
Donador hujayralning nozik, miyelinlanmagan aksonlari poʻstloqning yuqori (molekulyar) qavatiga vertical koʻtariladi, shu yerda ikkiga boʻlinadi va har biri gorizontal yoʻnalib parallel tolani hosil qiladi; vertikal shoxning ikki gorizontal tarmoqqa boʻlinishi ajralib turuvchi “T” shaklini beradi. Odam parallel tolasi boʻlinishdan keyin har bir tomonga oʻrtacha 3 mm, umumiy 6 mm (poʻstloq qavatning 1/10 qismi) tarqaladi.[9] Parallel tolalar oʻz yoʻlida Purkinye hujayralarining dendrit daraxtlarini kesib oʻtadi va har 3-5 tadan bittasi bilan bogʻlanib, Purkinye hujayralari denrit oʻsiqchalari bilan umumiy 80-100 ta sinaptik bogʻlanishlar hosil qiladi.[9] Donador hujayralar neyromediator sifatida glutamat kislotasini ishlatadi va shuning uchun nishonlariga qoʻzgʻatuvchi ta’sir qiladi.[9]
Donador hujayralar impulsni moxsimon tolalardan qabul qiladi, lekin son jihatdan ulardan ancha koʻp: 200 taga 1 ta (odamlarda). Shunday qilib donador hujayralar populyatsiyasi faollik holati moxsimon tolalardagi axborot bilan bir xil, lekin anchagina qimmat usulda koʻchiriladi. Donador hujayralar juda kichik va zich joylashgani uchun ularning spayk faolligini tajrivadagi hayvonlarda yozib olish qiyin, shuning uchun nazariyalarga asos sifatida foydalanish uchun ma’lumotlar kam. Ularning funksiyasi boʻyicha eng mashhur gʻoya 1969-yili David Marr tomonidan ilgari surilgan: ular moxsimon tolalardan kelgan impulslar kombinatsiyalarini shifrlaydilar. Bu, har bir donador hujayra 4-5 tagina moxsimon toladan impuls oladi va faqatgina biitasidan impuls kelganida javob bermaydi, balki bittadan koʻpidan kelganda javob beradi, degan fikrga asoslangan. Kombinatsiyalangan shifrlash sxemasi miyachaga, unga keladigan impulslarni faqatgina moxsimon tolalar olib keladiganga nisbatan nozikroq farqlash imkonini beradi.[24]
Moxsimon tolalar
[tahrir | manbasini tahrirlash]Moxsimon tolalar donador qavatga ularning hosil boʻlish nuqtalaridan kiradi, koʻpchiligi nucleus pontineusdan koʻtariladi, qolgan qismi orqa miya, vestibulyar yadrolar va boshqalardan keladi. Odam miyachasida moxsimon tolalarning umumiy soni taxminan 200 millionta.[9] Bu tolalar donador hujayralar va miyachaning chuqur yadrolari bilan qoʻzgʻatuvchi sinapslar hosil qiladi. Donador qavat ichida moxsimon tolalar “rozetkalar” deb ataluvchi kengaymalar hosil qiladi. Moxsimon tolalar va donador hujayra dendritlari orasidagi kontakt glomerula deb ataluvchi strukturalar ichida kechadi. Har bir glomerulada markazdagi moxsimon tola rozetkasi va unga bogʻlangan 20 tagacha donador hujayra dendrit changallari bor. Golji hujayralari terminallari strukturani infiltratlaydi va donador hujayra dendritlarida ingibirlovchi sinapslar hosil qiladi. Butun assembleya glial hujayralar gʻilofi bilan oʻralgan.[9] Har bir moxsimon tola bir qancha miyacha foliylariga kollateral tarmoqlar beradi va umumiy 20-30 ta rozetkalar hosil qiladi; shunday qilib bitta moxsimon tola taxminan 400-600 ta donador hujayralar bilan kontakt hosil qiladi.[9]
Koʻtariluvchi tolalar
[tahrir | manbasini tahrirlash]Purkinye hujayralari miya ustunining kontralateral qismidagi pastki oliva yadrosidan ham koʻtariluvchi tolalar orqali impuls qabul qiladi. Pastki oliva uzunchoq miyada joylashgan va orqa miya, miya ustuni va yarimsharlar poʻstlogʻidan impuls qabul qilib toʻgʻri miyachaga joʻnatadi. Koʻtariluvchi tola miyacha poʻstlogʻiga kirishdan oldin miyacha chuqur yadrolariga kollaterallar beradi. Miyacha poʻstlogʻida har biri 10 ta terminal tarmoqlarga boʻlinadi va ularning har biri alohida Purkinye tolasiga bogʻlanadi.[9] Taqqoslash uchun, har bir Purkinye hujayrasi 100 000dan ortiq parallel tolalardan va bir dona koʻtariluvchi toladan impuls qabul qiladi; lekin shu bir dona tola Purkinye hujayralarining dendritlariga “koʻtarilib” ularning atrofida oʻraladi va 300 tagacha sinapslar hosil qiladi.[9] Bu tarmoq shu qadar kuchliki, koʻtariluvchi toladagi birgina harakat potensiali Purkinye hujayrasida murakkab spayk hosil boʻlishiga olib keladi: toʻsatdan bir qancha spayklar qatori hosil boʻladi. Ular amplitudasi pasayib boradi va ulardan keyin faollik toʻxtagan pauza keladi. Koʻtariluvchi tola sinapslari hujayra tanasi va proksimal dendritlarni qoplab oladi; bu zonadan parallel tola impulslari kirmaydi.[9]
Koʻtariluvchi tolalar kam qoʻzgʻaladi, lekin birgina koʻtariluvchi tola harakat potesiali nishon Purkinye hujayrasida toʻsatdan koʻplab harakat potensiallari chaqiradi (murakkab spayk. Purkinye hujayrasiga keluvchi parallel va koʻtariluvchi tolalar farqi (100 000 dan ortiq va bir dona) miyacha anatomiyasining eng oʻziga xos xususiyati boʻlib, koʻplab nazariyalarga sababchi boʻlgan. Koʻtariluvchi tolalar funksiyasi miyachaga aloqador eng baxsli mavzudir. Hozirgi kunda buni tushuntiruvchi ikkita maktab bor: Marr va Albus maktabi gʻoyasi, koʻtariluvchi tola toʻgʻridan-toʻgʻri oʻrgatuvchi signallar tashiydi; boshqa gʻoya, miyachadan chiquvchi impulslarni bevosita bir shaklga keltirish. Ikkala qarash ham katta miqdordagi maqolalar bilan himoya qilingan. In the words of one review, "In trying to synthesize the various hypotheses on the function of the climbing fibers, one has the sense of looking at a drawing by Escher. Each point of view seems to account for a certain collection of findings, but when one attempts to put the different views together, a coherent picture of what the climbing fibers are doing does not appear. For the majority of researchers, the climbing fibers signal errors in motor performance, either in the usual manner of discharge frequency modulation or as a single announcement of an 'unexpected event'. For other investigators, the message lies in the degree of ensemble synchrony and rhythmicity among a population of climbing fibers."[21]
Chuqur yadrolar
[tahrir | manbasini tahrirlash]
Miyacha chuqur yadrolari miyacha oʻzagidagi oq modda ichida yotuvchi kulrang modda toʻplamlari. Ular vestibulyar yadrolarni hisobga olmaganda, miyachadan chiquvchi impulslar asosini tashkil qiladi. Bu yadrolar moxsimon va koʻtariluvchi tolalardan kollaterallar, shuningdek miyaha poʻstlogʻidagi Purkinye hujayralaridan ingibirlovchi impulslani qabul qiladi. Toʻrt yadro (tishsimon, sharsimon, probkasimon va choʻqqi), har biri bosh miya va miyacha poʻstlogʻining turli qismlari bilan bogʻlanadi. (Sharsimon va probkasimon yadrolar birgalikda aralashgan yadro (ing. interposed nucleus) deb ham ataladi). Choʻqqi va aralashgan yadrolar spinocerebellumga kiradi. Tishsimon yadro, sutemizuvchilarda boshqalarga nisbatan yirik boʻlib, nozik, ixcham kulrang modda qavatini hosil qiladi va miyacha poʻstlogʻining lateral qismlari bilan aloqa qiladi. Boʻlakcha-tuguncha (flocculonodular) boʻlakning boʻlakcha (flocculus) qismi miyacha poʻstlogʻidagi chuqur yadrolar bilan aloqa qilmaydigan yagona soha. U vestinulyar yadrolarga impuls beradi.[9]
Chuqur yadrolardagi koʻpchilik neyronlarda yirik hujayra tanalari va sferik dendrit daraxtlari (taxminan 400 μm radiusda) va neyromediator sifatida glutamat kislotani ishlatadi. Bu hujayralar miyacha tashqaridagi turli nishonlarga impuls beradi. Ular bilan aralash joylashgan kam sondagi kichik hujayralar neyromediator sifatida GAMKni ishlatadi va koʻtariluvchi tolalar chiquvchi pastki oliva yadrosiga impuls yuboradi. Shunday qilib, nukleo-olivar proyeksiya ingibitor ta’sir qiladi, koʻtariluvchi tolalar boʻlsa yadroga qoʻzgʻatuvchi ta’sir qiladi. Yadro hujayralarining kichik klasteri unga koʻtariluvchi tola joʻnatuvchi oliva hujayralari klasteriga impuld yuborishi haqida dalil mavjud; ikkala yoʻnalishda ham kuchli va mos keluvchi topografiya mavjud.[9]
Purkinye hujayrasi aksoni chuqur yadrolarga kirganda tarmoqlanadi va katta va kichik hujayralarga bogʻlanadi, lekin bogʻlanishlarning umumiy soni bor yoʻgʻi 35 ta (mushuklarda). Aksincha, birgina chuqur yadro hujayrasi taxminan 860 ta Purkinye hujayrasidan impuls qabul qiladi (yana mushuklarda).[9]
Boʻlimlari
[tahrir | manbasini tahrirlash]
Umumiy anatomik nuqtayi nazardan, miyacha poʻstlogʻi gomogen toʻqima massasidan iborat. Mikroanatomik nuqtayi nazardan ham bu massaning hamma qismlari oʻxshash ichki tuzilishga ega. Lekin, miyacha boʻlimlarga boʻlinganligi haqida koʻplab ma’lumotlar bor. Yirik boʻlimlar umumiy “zona” deb ataladi; ular yanada kichikroq boʻlimlarga boʻlinadi va “mikrozona” deb ataladi.[25]
Boʻlimli struktura haqidagi ilk tushunchalar miyacha poʻstlogʻining turli qismlaridagi hujayralarning reseptiv faoliyatini oʻrganish mobaynida paydo boʻldi.[25] Tananing har bir qismi miyachaning maxsus nuqtalarida akslanadi, lekin koʻplab marta takrorlanib akslanadi va “singan somatotopiya” deb ataluvchi holatda joylashadi.[26] Boʻlimlarga boʻlinish haqidagi yaqqolroq belgilar miyachadagi ma’lum turdagi oqsillarni immunoboʻyash orqali olindi. Eng mashhur markerlar “zebrinlar” deb ataladi, chunki ularni boʻyash zebra chiziqlarini eslatuvchi tasvirlar beradi. Zebrinlar va boʻlimlarga ajratuvchi boshqa markalar yordamida hosil qilingan chiziqlar miyacha pushtalariga perpendikulyar yoʻnalgan – ular mediolateral yoʻnalishda tor, uzun yoʻnalishda choʻzilgan. Turli markerlar turli xil chiziq toʻplamlarini hosil qiladi, joylashuviga koʻra funksiyasi boʻyicha kengligi va uzunligi farq qiladi, lekin ularning hammasi umumiy shaklga ega.[25]
1970-yillar oxirida Oskarsson poʻstloq zonalari kichikroq mikrozonalarga boʻlinishi mumkinligi haqida aytdi.[27] Mikrozona – bir xil somatotopic reseptiv sohaga ega boʻlgan Purkinye hujayralari guruhi. Mikrozonalar 1000 tagacha Purkinye hujayralari tutadi va poʻstloq pushtalariga perpendikulyar joylashgan uzun, tor chiziqlarda joylashgan.[25] Diagrammada koʻrsatilganidek, Purkinye hujayralari dendritlari mikrozona bilan bir yoʻnalishda joylashgan, parallel tolalar boʻlsa toʻgʻri burchak ostida kesib oʻtadi.[9]
Mikrozona tuzilishini faqatgina reseptiv soha tavsiflamaydi: pastki oliva yadrosidan chiquvchi koʻtariluvchi tolalar ham teng ahamiyatli. Koʻtariluvchi tolaning tarmoqlari (odatda 10 tagacha raqamlanadi) odatda bir mikrozonadagi Purkinye hujayralarini faollashtiradi. Bir mikrozonaga koʻtariluvchi tolalar yuboruvchi oliva neyronlari tirqishli tutashmalar orqali birlashgan va bu ular faolligini sinxronlashtirib, mikrozonadagi Purkinye hujayralarida millisekundlik vaqt davomida korrelyatsiyalangan murakkab spayklar faolligiga olib keladi.[25] Shuningdek, mikrozonaga kiruvchi Purkinye hujayralari hammasi aksonlarini miyacha chuqur yadrolaridagi oʻxshash hujayra klasterlariga yuboradi.[25] Savatsimon hujayralar aksonlari uzunlik yoʻnalishida mediolateral yoʻnalishga qaraganda birmuncha uzunroq va bu ularni bitta mikrozonaga kattalik qilishiga sabab boʻladi.[25] Bu tuzilmalardagi tartib mikrozona ichidagi hujayralarning aloqalari mikrozonalararo aloqalardan koʻra birmuncha kuchliroqligiga olib keladi.[25]
2005-yilda Richard Apps va Martin Garwicz mikrozonalar yirikroq multizonal mikrokomplekslar tarkibiga kirishi haqidagi ma’lumotlarni umulashtirdilar. Bunday mikrokompleks bir miyacha chuqur neyronlari guruhiga akslanuvchi alohida poʻstloq mikrozonalaridan tashkil topgan. Shungdek mos oliva neyronlari guruhlari shu mikrokompleksga kiruvchi barcha mikrozonalarga va chuqur yadrolarga akslanadi,.[25]
Qon bilan ta’minlanishi
[tahrir | manbasini tahrirlash]Miyacha uch juft asosiy arteriyalardan qon oladi: Yuqorigi miyacha arteriyasi, oldingi pastki miyacha arteriyasi, orqa pastki miyacha arteriyasi. Yuqorigi arteriya miyachaning yuqorigi sohalarini qon bilan ta’minlaydi. U yuqorigi yuzaga yoʻnaladi va yumshoq pardada oldingi va orqa pastki miyacha arteriyalari bilan anastomozlar hosil qiluvchi tarmoqlar beradi. Oldingi pastki miyacha arteriyasi miyachaning pastki yuzasining oldingi qismini qon bilan ta’minlaydi. Orqa pastki miyacha arteriyasi pastki yuzaga kelganda medial va lateral tarmoqlarga ajraladi. Medial tarmoq miyacha yarimsharlari orasidagi kertik sohaga davom etadi; lateral tarmoq boʻlsamiyachaning pastki yuzasini, asosan lateral qirgʻoqlarini qon bilan ta’minlab, oldingi pastki va yuqorigi arteriyalar bilan anastomozlar hosil qiladi.
Funksiyasi
[tahrir | manbasini tahrirlash]Miyacha fuksiyasi boʻyicha muhim ma’lumotlar uning jarohati oqibatlarini oʻrganish orqali olingan. Miyacha disfunksiyasi bor hayvonlar va odamlarda, eng avvalo tananing miyachaning zaralangan qismi bilan bir tomonida harakat boshqariluvi bilan muammolar kuzatiladi. Ular umumiy harakatlarni amalga oshirishda davom etadi, lekin aniqlik yoʻqolgan, harakatlari qaltiroqli, koordinatsiyalanmagan yoki vaqti notoʻgʻri. Miyacha funksiyasi uchun standart test qoʻl uzunligidagi nishonga barmoq uchini tegizish: sogʻlom odam barmogʻini tez va toʻgʻri trayektoriyada harakatlantiradi, miyacha jarohati bor inson boʻlsa sekin va qaltirab, bir qancha toʻgʻrilanishlar bilan erishadi. Harakatga bogʻliq boʻlmagan funksiyalardagi yetishmovchiliklarni aniqlash ancha qiyin. Shunday qilib, oʻn yillar oldingi xulosaga koʻra miyachaning asosiy funksiyasi harakatni kalibratsiya qilish, harakatni boshlash yoki qaysi harakatlar bajarilishini hal qilish emas.[11]
1990-yillarda miyachaning funksiyasi faqat harakatga bogʻliq deb hisoblangan, lekin yangi izlanishlar bu qarashni soʻroq ostiga qoʻydi. Funksional tomografiyada oʻrganishlar til, diqqat, hayoliy tasavvur qilish paytida miyacha faollashuvini koʻrsatdi; korrelyatsiyani oʻrganish miyacha va yarimsharlar poʻstlogʻining harakatga bogʻliq boʻlmagan sohalari orasidagi aloqalarni koʻrsatdi; shuningdek miyacha zararlanishiga ega odamlarda harakatga bogʻliq boʻlmagan turlicha simptomlar kuzatildi.[28][29] Xususan, miyacha kognitiv affektiv sindromi yoki Schmahmann sindromi[30] kattalarda[31] va bolalarda tasvirlangan.[32] Miyachaning funksional MRT yordamida funksional xaritalanishi miyacha poʻstlogʻining yarmidan koʻpi yarimsharlar poʻstlogʻining assosiativ zonalari bilan aloqasi borligini taxmin qilish imkonini berdi.[33]
Kenji Doya miyachaning funksiyasini harakat-holatga ta’siri boʻyicha emas, balki bajaradigan nevrologik hisoblashlari orqali yaxshiroq tushuntirish mumkin deb hisoblaydi; miyacha juda koʻp miqdordagi koʻproq yoki kamroq mustaqil modullardan iborat, hammasining ichki tuzilishi oʻxshash geometric tartibda va taxminlarga koʻra bir xil hisoblashlarni amalga oshiradi. Agarda modulning kiruvchi va chiquvchi bogʻlabishlari harakat zonalari bilan boʻlsa (koʻpchiligida shunaqa), unda modul harakatda ishtirok etadi; ammo, agarda bogʻlanishlar harakatga bogʻliq boʻlmagan sohalar bilan boʻlsa, modul ham holatning harakatsiz turlarini korrelyatsiya qilishi koʻrinadi. Shunday qilib miyacha his-tuygʻu, harakat va holat kabi funksional xususiyatlar boshqariluviga jalb qilingan.[34][35] Doya fikriga koʻra, miyachani muhitning "ichki modellari" asosida harakatni oldindan bashorat qilib tanlash yoki nazoratli oʻrganish moslamasi sifatida tushunish eng toʻgʻri. Undan farqli, basal ganglia mustahkamlovchi oʻrganishni amalga oshirsa, yarimsharlar poʻstlogʻi nazoratsiz oʻrganishni amalga oshiradi.[29][36]
Prinsiplari
[tahrir | manbasini tahrirlash]Miyacha anatomiyasining nisbatan oddiyligi va tartibliligi erta davrlarda uning hisoblash funksiyasi ham shunday oddiy boʻlsa kerak degan taxmin qilishga asos boʻlgan. Bu miyacha elektrofiziologiyasi haqidagi ilk kitoblardan boʻlgan John C. Eccles, Masao Ito va János Szentágothaining Miyacha neyronal mashina sifatida asarida ham keltirilgan.[37] Garchi miyacha funksiyasini toʻliq tushunish qiyinligicha qolayotgan boʻlsada, kamida toʻrtta muhim prinsiplar mavjud: (1) feedforward processing (orada qayta ishlash), (2) divergensiya va konvergensiya, (3) modullilik va (4) plastiklik.
- Feedforward processing: miyacha bosh miyaning boshqa qismlaridan (xususan yarimsharlar poʻstlogʻidan) signalni qayta ishlash deyarli toʻliq feedforward usulida kechishi bilan farq qiladi. Feedforward – signallarning kirishdan chiqishga toʻgʻridan-toʻgʻri uzatilmay, juda kichik davriy ichki uzatmalar bilan oʻtishi. Mavjud boʻlgan kichik miqdordagi qaytish umumiy ingibirlovchidan iborat; umumiy qoʻzgʻatuvchi halqalar mavjud emas. Miyachaning feedforward tartibida ishlashi, yarimsharlar poʻstlogʻidan farqli ravishda, oʻz-oʻzini quvvatlovchi neyral faollik hosil qila olmasligini bildiradi. Halqaga kirgan signallar tartib bilan qayta ishlanadi va chiqib ketadi. Eccles, Ito va Szentágothai yozganidak, “Neyronal qoʻzgʻalishning reverberator zanjirlaridagi ushbu olib tashlashlar, miyachani kompyuter kabi ishlashida shubxasiz katta afzallik beradi. Chunki, taxminan nerv sistemasining qolgan qismlari miyachadan talab qiluvchi narsa kompleks reverberator halqalarni chiqarish emas, balki ma’lum informatsiya toʻplamiga tez va aniq javob berishdir."[38]
- Divergensiya va konvergensiya: odam miyachasida, 200 millon moxsimon tolalardan kelgan axborot 40 milliard donador hujayralariga tarqatiladi, ularning parallel tolalari 15 million Purkinye hujayralarida kesishadi.[9] Uzunlik chizigʻI hosil qilib joylashganligi uchun, mikrozonaga kiruvchi 1000 ta Purkinye hujayrasi 100 milliontagacha parallel tolalardan impuls oladi va 50 dan kamroq chuqur yadroning hujayralari guruhiga yuboradi.[25] shunday qilib, miyacha tarmogʻi kam miqdorda impulslar qabul qilib, ularni juda tartibli tuzilgan ichki tarmoq orqali kengaytirib qayta ishlaydi va natijani juda kam sondagi hujayralar orqali tashqariga yuboradi.
- Modullilik: Miyacha tizimi funnksional jihatdan yuzlab, minglab koʻp yoki kam darajadagi mustaqillikka ega modullarga boʻlinadi. Barcha modular oʻxshash ichki tuzilishga ega, lekin kirish va chiqish turlicha. Modul (Apps va Garwicz terminologiyasi boʻyicha multizonal mikrokompartment) pastki oliva yadrosidagi neyronlarning kichik klasteri, miyacha poʻstlogʻidagi Purkinye hujayralarining uzun, tor chiziqlari toʻplami (mikrozonalar) va miyacha chuqur yadrolaining biridagi neyronlarning kichik klasteridan tashkil topgan. Turli modular kiruvchi impulslarni moxsimon va parallel tolalar orqali almashishi mumkin, lekin qolgan sohalardagi funksiyalari mustaqil – bir moduldan chiquvchi impuls boshqa modullarga bevosita ta’siq qilmaydi.[25]
- Plastiklik: Purkinye hujayralari bilan parallel tolalar orasidagi va moxsimon tolalar bilan chuqur yadro hujayralari orasidagi sinapslar ularning kuchi oʻxgarishiga sezgir. Bitta miyacha modulida, milliard parallel tolalardan kelgan impuls 50 dan kamroq chuqur yadro neyronlarida kesishadi va har bir parallel tolaning shu yadro hujayralariga ta’siri tartiblangan. Bu tartib miyacha kiruvchi va chiquvchi impulslari orasidagi munosabatni puxta sozlash uchun ulkan moslanuvchanlik beradi.[39]
Oʻrganish
[tahrir | manbasini tahrirlash]Miyacha harakat oʻrganishning ma’lum turlarida muhim rol oʻynashi haqida muhim dalillar mavjud. Miyacha aniq qatnashadigan topshiriqlar, yuqori tartibni talab qiluvchi harakatlardir. Bu masalada bir qancha munozaralar mavjud ya’ni oʻrganish miyachaning oʻzida kechadimi yoki miyaning boshqa qismlarida oʻrganishga sabab boʻluvchi signallar yuboradimi.[39] Oʻrganish miyachada kechishi haqidagi koʻplab nazariyalar David Marr[24] va James Albuslarning,[6] gʻoyalari asosida yaratilgan. Ular koʻtariluvchi tolalar parallel tola – Purkinye hujayrasi sinapsida modifikatsiyaga sabab boʻluvchi oʻrgatuvchi signal tashiydi degan postulatni ilgari surdilar.[40] Marr taxminiga koʻra koʻtariluvchi toladan keluvchi impuls sinxron qoʻzgʻalgan parallel tola impulsini kuchaytiradi. Keying koʻplab miyachada oʻrganish modellari, koʻtariluvchi tola faolligi xato signal boʻlib, sinxron faollangan parallel tola impulslarini kuchsizlantiradi deb taxmin qiluvchi Albusga ergashgan. Bu keyingi modellardan ba’zilari, masalan Fujitaning Adaptiv Filtr modeli[41] miyacha funksiyasini optimal boshqaruv nazariyasi bilan tushuntirishga harakat qiladi.
Koʻtariluvchi tola faolligi xato signal sifatida ta’sir qilishi haqidagi gʻoya koʻplab tajribalarda sinab koʻrildi, ularning ba’zilari qoʻllab-quvvatlasa, qolgani gumonlarni kuchaytiradi.[21] Gilbert va Thachning 1977-yilda sohadagi boshlangʻich tajribalarida, maymunlarning Purkinye hujayralari topshiriqni bajarishni oʻrganishda ijro darajasi past boʻlgan davrlarda kuchaygan murakkab spayk – koʻtariluvchi tolalar faooligining ishonchli koʻrsatkichi – faolligini koʻrsatdi.[42] Mushuklardagi harakat oʻrganish boʻyicha bir qancha tadqiqotlar yoʻnaltirilgan harakat va uni cheklovchi harakatlar birga kelgan vaqtda murakkab spayk faolligini koʻrsatdi. vestibulo-okulyar refleks (bosh burilganda vizual tasvirni retinada ushlab turuvchi)ni tadqiq qilish koʻtariluvchi tola “retinal siljish”ni chaqirgan, bevosita boʻlmagan yoʻl bilan.[21]
Miyachada oʻrganish topshiriqlaridan keng oʻrganilganlaridan biri koʻz pirpirashini chiniqtirish(ing. eyeblink conditioning) paradigmasi boʻlib, tovush yoki yorugʻlik kabi neytral shartli ta’sir, koʻz pirpirashiga sabab boʻluvchi havo pudash kabi shartsiz ta’sir bilan juftlashtiriladi. Bunday shartli va shartsiz ta’sirlar bir necha marta qaytarilganidan soʻng, shartli ta’sir shartsiz ta’sir boʻlmasidan oldin koʻz yumilishini chaqiradi, shartli javob. Tajribalar koʻrsatishicha interpozitsiyalangan yadro (miyacha chuqur yadrolaridan biri)ning ma’lum qismi yoki miyacha poʻstlogʻining ma’lum nuqtalaridagi shikastlanishlar chiniqtirilgan koʻz yumish javobini toʻxtatadi. Agar miyachadan chiquvchi impulslar farmakologik inaktivatsiya qilinsa va miyacha ichi yoʻllari butun boʻlsa, hayvon javobni koʻrsata olmasa ham oʻrganish sodir boʻladi, aksincha miyacha ichi yoʻllari zararlangan boʻlsa oʻrganish kuzatilmaydi – bu faktlar birgalikda olinganda, oʻrganish miyacha ichida kechishiga kuchli misol boʻladi.[43]
Nazariyalar va hisoblash modellari
[tahrir | manbasini tahrirlash]
Miyacha anatomik tuzilishi va funksiyalarini oʻrganishdagi ulkan baza, uni nazariya uchun unumdor maydonga aylantirdi – miyacha funksiyasi boʻyicha yaratilgan nazariyalar miyaning boshqa har qanday qismidagiga nisbatan koʻpdir, balki. Ularning ichida eng farqlilari “oʻrganish nazariyalari” va “ijro nazariyalari” – ular, miyachaning oʻrganishdagi rolini hisobga olish uchun sinaptik plastiklikdan foydalanuvchi nazariyalar va aksincha miyachadagi signal prosessingi asosidagi davom etayotgan harakat-holatning xususiyatlarini hisobga oluvchi nazariyalar. Ikkala turdagi bir qancha nazariyalar matematik modellar sifatida ishlab chiqilgan va kompyuter yordamida simulyatsiya qilingan.[40]
Ehtimol eng birlamchi “ijro” nazariyasi Valentino Braitenbergning “kechikkan chiziq” gipotezasi boʻlgandir. Original nazariya 1958-yilda Braitenberg va Roger Atwood tomonidan ilgari surilgan, unga koʻra parallel tolalardan signalning sekin tarqatilishi kechikishga sabab boʻladi va miyachaga ma’lum darchalarda vaqt munosabatlarini sezishga yordam beradi.[44] Tajriba natijalari nazariyaning original variantini tasdiqlamadi, lekin Braitenberg modifikatsiyalangan versiyalar bilan bahslashishda davom etdi.[45] Miyacha vaqt sistemasi sifatida ishlashi haqidagi gipotezani Richard Ivry ham qoʻllab quvvatlagan.[46] Boshqa nufuzli “ijro” nazariyasi Pellionisz va Llináslarning Tenzor tarmoq nazariyasi boʻlib, miyacha hisoblashlari sezgini harakat koordinatsiyalariga aylantirish gʻoyasini yuqori darajada matematik qayta ishlashgan.[47]
“Oʻrganish” kategoriyasining deyarli barcha nazariyalari Marr va Albus maqolalaridan olingan. Marrning 1969-yildagi maqolasiga koʻra, miyacha oʻrganish qurilmasi boʻlib, koʻtariluvchi tolalardagi elementar harakat impulslari bilan moxsimon tolalardagi sezgi impulslarini assotsiatsiyalaydi.[24] Albus 1971-yilda miyacha Purkinye hujayralari perseptron kabi ishlashini, neyral ilhomlangan abstract oʻrganish mashinasi ekanligini ta’kidlaydi.[6] Marr va Albus nazariyalari orasidagi asosiy farq, Marr koʻtariluvchi tola faolligi parallel tola sinapsi kuchayishiga sabab boʻladi desa, Albus kuchsizlantiradi deb hisoblaydi. Albus oʻz versiyasining Artikulyatsiya boshqaruvining miyacha modeli (CMAC (Cerebellar Model Articulation Controller)) deb nomlangan dasturiy algoritmini ham ishlab chiqdi va bu koʻplab ilovalarda sinab koʻrildi.[48]
Klinik ahamiyati
[tahrir | manbasini tahrirlash]

Miyacha zararlanishi odatda harakatga bogʻliq simptomlarga sabab boʻladi, detallari miyachaning qaysi qismi va qanday zararlanganiga bogʻliq. Flocculonodular boʻlakning zararlanishi muvozanat yoʻqolishi va qisman buzilgan, tartibsiz qadam tashlash, keng doirada balans qilishning qiyinlashuvi bilan namoyon boʻladi.[11] Lateral zonaning zararlanishi odatda ihtiyoriy va rejali, malakali harakatlarda muammolarga sabab boʻlib, harakatning kuchi, yoʻnalishi, tezligi va amplitudasida xatoliklarga sabab boʻladi. Boshqa koʻrinishlarga kiradi, gipotoniya (pasaygan muskul tonusi), disartriya [[gapirish artikulyatsiyasi bilan muammolar), dismetriya (masofani baholash yoki harakat chegaralarida muammolar), disdiadoxokineziya (piyoda yurish kabi tez almashib turuvchi harakatlarni bajara olmaslik), buzilgan nazorat refleksi yoki rebound fenomeni, maqsadli qaltirash (ing. intention tremor) (qarama-qarshi muskul guruhlarining almashib turuvchi qisqarishlari natijasida yuzaga kelgan noixtiyoriy harakatlar).[49][50] Oʻrta chiziq sohasidagi zararlanish butun tana harakatlarini buzadi, lateralroq joylashgan jarohatlar boʻlsa, oyoq-qoʻllarning nozik harakatlarini buzadi. Miyachaning yuqori qismi jarohati qadam tashlash buzilishi va oyoq koordinatsiyalaridagi muammolarni chaqirsa, pastki qismi jarohati panja va qoʻlllarning koordinatsiyalashmagan va maqsadga kam yoʻnaltirilgan harakatlarga, tez bajarishdagi qiyinchiliklarga sabab boʻladi.[11] Bu harakat simptomlari kompleksi ataksiya deb nomlanadi.
Miyacha muammolarini aniqlash uchun nevrologik tekshiruvlarga qadamni baholash (keng asosli qadam ataksiya belgisi), barmoq bilan tegish testlari va tana holatini baholash kiradi.[4] Miyacha disfunksiyasi aniqlanganda, boʻlishi mumkin boʻlgan har qanday stuktura buzilishlarini batafsil tasvirini olish uchun magnit rezonans tomografiya oʻtkazilishi mumkin.[51]
Miyaza zararlanishiga sabab boʻluvchi tibbiy muammolar roʻyhati uzun: insult, gemorragiya, miya shishi, oʻsmalar, alkogolizm, oʻq tegish yoki portlashdan keying jismoniy jarohatlar va olivopontocerebellar atrofiya kabi surunkali degenerativ holatlar.[52][53] Migren bosh ogʻrigʻining ba’zi shakllari ham turli darajadagi vaqtinchalik miyacha disfunksiyasini chaqirishi mumkin.[54] Miyacha zararlanishiga prion kasalligi[55] va Guillain–Barré sindromining varianti boʻlgan Miller Fisher sindromi kabi infeksion holatlar ham sabab boʻlishi mumkin.
Yoshga bogʻliq xususiyatlari
[tahrir | manbasini tahrirlash]Odam miyachasi yoshga bogʻliq holatda oʻzgaradi. Bu oʻzgarishlar miyaning boshqa qismlaridan farq qilishi mumkin. Yuz yoshdan oshganlarda, epigenetik soat sifatida ma’lum boʻlgan toʻqima yoshi epigenetik biomarkeriga koʻra, miyacha miyaning eng yosh sohasi (va tana qismi): yuz yoshlilarda taxminan qilinganidan taxminan 15 yilga yoshroq.[56] Keyinchalik, gen ekspressiyasi ma’lumotlari odam miyachasida yarimsharlar poʻstlogʻiga nisbatan kam yoshga bogʻliq zararlanishlar borligini koʻrsatdi.[57] Ba’zi tadqiqotlar hujayralar soni yoki toʻqima hajmi kamayishini koʻrsatgan, lekin bu masaladagi ma’lumotlar koʻp emas.[58][59]
Rivojlanishdagi va degenerativ buzilishlar
[tahrir | manbasini tahrirlash]
Tugʻma yetishmovchiliklar, irsiy buzilishlar va orttirilgan holat miyacha tuzilishiga, shuningdek miyacha funksiyasiga ta’sir qilishi mumkin. Sabab boʻluvchi holatni bartaraf qilib boʻlmasa, yagona davo chorasi odamlarga shu muammolar bilan yashashga yordam berishdir.[60] Homiladorlikning 18-20 haftalarida homila miyachasini ultratovush skaneri orqali vizualizatsiya qilish homila nerv naychasi defektlari skriningida 99% lik sezgirlikka ega.[61]
Normal rivojlanishda, endogen sonic hedgehog signallari tashqi donador qavatdagi (TDQ) miyacha donador neyronlari progenitorlarining (MDNP) tez proliferatsiyasini stimullaydi. Miyacha rivojlanishi kech embriogenez va erta postnatal davrda kichadi, erta rivojlanishdagi MDNPning TDQ tepaligidagi proliferatsiyasi bilan (sichqonda 7-postnatal kun).[62] MDNPlar oxirida miyacha donador hujayralari (miyacha granulyar neyronlari, MGN, deb ham ataladi)ga differensiatsiyalanib, ichki donador qavatga koʻchadi va yetuk miyachani hosil qiladi (sichqonda 20-postnatal kun).[62] Sonic hedgehog signalingni anormal faollashuviga sabab boʻluvchi mutatsiyalar Gorlin sindromi bor odamlarda va genetik injenerlangan sichqon modellarida miyacha oʻsmasi (medulloblastoma)ga nisbatan moyillikka sabab boʻladi.[63][64]
Miyacha chuvalchangsimon qismining tugʻma majruhligi yoki toʻliq rivojlanmasligi (gipoplaziya) Dandy-Walker sindromi va Joubert sindromining ikkovi uchun ham xos xususiyat.[65][66] Juda kamdan-kam holatlarda miyacha umuman boʻlmasligi mumkin.[67] Irsiy nevrologik kasalliklar boʻlgan Machado-Joseph kasalligi, ataksiya teleangiektaziya va Friedreich ataksiyasi miyacha yoʻqolishi bilan bogʻliq progressiv neyrodegeneratsiyaga sabab boʻladi.[52][60] Oʻz navbatida, miyachadan tashqaridagi tugʻma miya majruhligi miyacha toʻqimasining churrasiga sabab boʻladi va Arnold-Chiari majruhligining ba’zi shakllarida koʻrinadi.[68]
Miyacha degeneratsiyasi bilan bogʻlangan boshqa holatlarga idiopatik progressiv nevrologik buzilishlar boʻlgan koʻplab sistemalar atrofiyasi va Ramsay Hunt sindromi I tip,[69][70] va avtoimmun buzilish boʻlgan paraneoplastik miyacha degeneratsiyasi kiradi, bunda tananing boshqa qismida joylashgan oʻsma miyachada neyronlar yoʻqolishiga olib keluvchi avtoimmun javobni chaqiradi.[71] Miyacha atrofiyasiga beriberi va Wernicke-Korsakoff sindromida kuzatiluvchi oʻtkir vitamin B1 (tiamin) yetishmovchiligi[72] yoki vitamin E yetishmovchiligi ham sabab boʻlishi mumkin[60]
Miyacha atrofiyasi boshqa koʻplab nevrologik buzilishlar: Huntington kasalligi, tarqalgan skleroz,[55] essensial titrash, progressiv mioklonik epilepsiya va Niemann–Pick kasalligida ham oʻrganilgan. Miyacha atrofiyasi ogʻir metallar yoki farmasevtik yoki tinchlantiruvchi dorilar kabi toksinlardan zaharlanish natijasida ham yuzaga kelishi mumkin.[60]
Ogʻriq
[tahrir | manbasini tahrirlash]Miyacha ogʻriq shakllanishida qatnashadi degan umumiy konsensus mavjud.[73][74] Miyacha tushuvchi cortico-cerebellar yoʻllardan ham, koʻtariluvchi spino-cerebellar yoʻllardan ham, nucleus pontineus va pastki oliva yadrolari orqali ogʻriq impulslarini qabul qiladi. Bu informatsiyaning bir qismi harakat sistemasiga oʻtkaziladi va ogʻriq intensivligiga qarab harakatdagi ogʻriqdan qochishni chaqiradi.
Bu bevosita va bilvosita ogʻriq impulslari, uzoq muddatli ogʻriqdan qochish harakatiga sabab boʻladi deb hisoblanadi. Natijada surunkali gavdani tutishdagi oʻzgarishlar va oʻz navbatida vestibulyar va proprioseptiv yadrolarning anatomik va funksional oʻzgarishi yuzaga keladi. Shunday qilib, surunkali neyropatik ogʻriq hindbrainda, shuningdek miyachada makroskopik anatomik qayta modellanishni chaqirishi mumkin.[23] Bu qayta modellanishning magnitudasi va neyron progenitor markerlarini induksiya qilish bu oʻzgarishlarda voyaga yetganlar neyrogenezining hissasi borligini koʻrsatadi.
Solishtirma anatomiya va evolyutsiya
[tahrir | manbasini tahrirlash]
Miyacha yoʻllari umurtqalilarning barcha sinflarida: baliqlar, sudralib yuruvchilar, qushlar va sutemizuvchilarda oʻxshash.[75] Shuningdek yaxshi rivojlangan bosh miyaga ega boshoyoqlilar, masalan sakkizoyoqlar analogik miya tuzilishiga ega.[76] Miyacha bosh miyaga ega barcha hayvon turlarida muhim funksiyalar bajarishi haqida dalillar mavjud.
Har xil umurtqalilar turlarida miyacha oʻlchami va shakli farq qiladi. Amfibiyalarda u kam rivojlangan, lampreylar va hagfishda miyachani miya ustunidan zoʻrgʻa ajratvolish mumkin. Garchi bu guruhlarda spinocerebellum mavjud boʻlsada, birlamchi strukturalar kichik, juft yadrolar vestibulocerebellumga mos keladi.[77] Miyacha reptiliyalarda yirikroq, qushlarda yanayam yirikroq, sutemizuvchilarda undanam yirik. Odamlardagi yirik juft va ixcham boʻlaklar sutemizuvchilar uchun xos, lekin umuman, miyacha boshqa guruhlarda yagona medial boʻlak va silliq yoki biroz burmalardan iborat. Sutemizuvchilarda neocerebellum miyachaning massa boʻyicha eng katta qismi, lekin boshqa umurtqalilarga spinocerebellum xos.[77]
Togʻayli baliqlar va suyakli baliqlarda miyacha odatdan tashqari yirik va murakkab. Muhim xususiyati, u ichki tuzilishi bilan sutemizuvchilar miyachasidan farq qiladi: baliq miyachasi ajralgan miyacha chuqur yadrolari tutmaydi. Aksincha, Purkinye hujayralarining birlamchi nishoni miyacha poʻstlogʻi boʻylab tarqalgan maxsus hujayralar boʻlib, ular sutemizuvchilarda yoʻq. Mormyrid baliqlari (kuchsiz elektrosensitiv chuchuk suv baliqlari oilasi)da miyacha qolgan miya qismlari birga olinganidan yirikroq. Uning eng katta qismi valvula deb ataluvchi maxsus struktura boʻlib, noodatiy tartibdagi arxitekturasi mavjud va asosiy impulslarni elektrosensor sistemadan qabul qiladi.[78]
Sutemizuvchilar miyachasining oʻziga xos xususiyati lateral boʻlaklarning kengayganligi boʻlib, asosiy aloqalari neokorteks bilan boʻladi. Maymunlar yirik dumsiz maymunlarga qarab rivojlanib borgan sari, lateral tolalarning kengayishi davom etib, oʻz navbatida neokorteksning frontal boʻlaklari kengayishi bilan kechadi. Qadimgi maymunsimon odam (hominid)lar va Homo sapiensda Pleistocene davri oʻrtalarigacha miyacha kengayishda davom etdi, lekin frontal boʻlaklar ancha tez kengaydi. Odam evolyutsiyasining eng yaqin davrlari, miyacha nisbiy oʻlchamlarining oshishi bilan bogʻliq boʻlishi mumkin, miyacha kengayib borishi bilan neokorteks oʻz oʻlchamlarini qandaydir ravishda kamaytirgan.[79] odam miyachasi oʻlchamlari, miyaning boshqa qismlariga nisbatan olib qaralganda, katta yarimsharlar oʻlchami kamayishi bilan kattalashib bormoqda.[80] Harakat topshiriqlari rivojlanishi va bajarilishi, jismlarning samoviy holatini aniqlay olish qobiliyati va oʻrganishninf miyachad kechishi, miyachaning oʻsishi odamning katta kognitiv qobiliyatlari bilan qaysidir darajada korrelyatsiyaga ega ekanligi haqida oʻylashga sabab boʻladi.[81] Miyachaning lateral yarimsharlari hozirgi kunda, odam va odamsimon maymunlarda qolgan maymunlarga nisbatan 2,7 marta kattaroq.[80] Miyacha oʻlchamidagi bu oʻzgarishlarni muskul massasining kattaligi bilan tushuntirib boʻlmaydi. Bu miyacha rivojlanishi miyaning boshqa qismlari bilan qalin bogʻlanganligi yoki miyachada kechuvchi neyral faoliyatlar yirik odamsimon maymunlar (Hominidae) evolyutsiyasida muhim boʻlganligini koʻrsatadi. Miyachaning kognitiv funksiyalardagi roliga koʻra, uning oʻlchamlari oshishi kognitiv kengayishda rol oʻynagan boʻlishi mumkin.[80]
Miyachaga oʻxshash strukturalar
[tahrir | manbasini tahrirlash]Koʻp umurtqalilar turlarida, miyacha va bir yoki koʻp miyachaga oʻxshash strukturalar, sitoarxitektonika va neyrokimyoviy jihatdan miyachani eslatuvchi bosh miya sohalari, mavjud.[75] Sutemizuvchilardan topilgan yagona miyachaga oʻxshash struktura orqa koxlear yadro (OKY) boʻlib, toʻgʻridan-toʻgʻri eshituv nervidan impuls qabul qiluvchi ikkita birlamchi sezuv yadrosining biridir. OKY qavatli struktura boʻlib, pastki qavatni tashkil qiluvchi donador hujayralar miyachadagi kabi, yuza qavatga boruvchi va unda gorizontal tarqaluvchi parallel tolalarni beradi. Yuza qavat GAMKergik cartwheel hujayralar tizimidan tashkil topgan boʻlib, ular anatomik va kimyoviy jihatdan Purkinye hujayralarga oʻxshash – ular parallel tolalardan impuls qabul qiladi, lekin koʻtariluvchi tolalarga oʻxshash strukturalar yoq. OKYning tashqariga impuls chiqaruvchi neyronlari piramidasimon hujayralardir. Ular glutamatergik, lekin ba’zi jihatlari bilan Purkinye tolalariga ham oʻxshaydi – ularda tikanaksimon, tekislangan yuza dendritlari parallel tolalardan impuls qabul qiladi, lekin ularda shuningdek eshituv nervi tolalaridan impuls qabul qiluvchi bazal dendritlari ham mavjud boʻlib, ular OKYda parallel tolalarga nisbatan toʻgʻri burchak ostida joylashadi. OKY kemiruvchilar va boshqa kichik hayviblarda yaxshi rivojlangan boʻlib va primatlarda anchagina kichraygan. Uning funksiyasi yaxshi oʻrganilmagan; eng mashhur taxmin tovush yoʻnalishini aniqlash bilan bogʻliq.[82]
Koʻplab baliq va amfibiya turlarida suvdagi toʻlqinlar bosimini sezuvchi yon chiziq sistemasi mavjud. Yon chiziq organidan birlamchi impuls qabul qiluvchi miya qismlaridan biri, medial oktavolateral yadro, miyachaga oʻxshash tuzilishga ega boʻlibm donador hujayralari va parallel tolalari bor. Elektr sezuvchi baliqlarda elektrosensor sitemadan impuls dorsal oktavolateral yadroga boradi, u ham miyachaga oʻxshash strukturaga ega. Ray-finned baliqlari koʻruv doʻmbogʻida miyachaga oʻxshash qavat – marginal qavat mavjud.[75]
Ushbu barcha miyachaga oʻxshash strukturalar sezgi bilan bogʻlangan, harakat bilan emas. Ularning barchasida Purkinyega oʻxshash neyronlar bilan modifikatsiyalanuvchi sinaps hosil qiluvchi parallel tolalarni beradigan donador hujayralar mavjud, lekin miyachadan farqli birontasida koʻtariluvchi tolalar yoʻq – buning oʻrniga ular periferik sezgi organlaridan toʻgʻridan-toʻgʻri impuls qabul qiladi. Ularning funksiyalari aniqlanmagan, lekin eng ishonchli taxmin, ular sezgi impulslarini qandaydir murakkab yoʻl bilan qayta ishlaydi va bu ehtimol tana holati oʻzgarishlarini kompensatsiya qilish uchundir.[75] Aslini olganda, James M. Bower va boshqalar qisman bu strukturalar va qisman miyacha ustidagi tadqiqotlar asosida miyacha oʻzi fundamental jihatdan sensor struktura degan fikrni beradilar: uning harakat boshqaruvi sezgi signallari asosida tanani qoʻzgʻatishi orqali amalga oshadi.[83] Bowerning fikrlariga qaramay, miyacha sutemizuvchilar harakatiga bevosita ta’sir qilishi haqida kuchli dalillar mavjud.[84][85]
Tarixi
[tahrir | manbasini tahrirlash]
Tavsiflar
[tahrir | manbasini tahrirlash]Hattoki ilk anatomlar ham, miyachani oʻziga xos qiyofasi tufayli farqlay olganlar. Aristotel va Herophilus (Galendan iqtibos) παρεγκεφαλίς (paregkephalis) deb atashgan, bosh miyani oʻzini boʻlsa ἐγκέφαλος (egkephalos) deb atashgan. Galenning kengaytirilgan tavsifi mavjudlarning eng qadimgisi. U miyachani harakat nervlarining manbasi deb taxmin qilgan.[86]
Keyingi koʻzga koʻrinarli rivojlanishlar Renessansgacha kuzatilmagan. Vezaliy miyachani qisqacha ta’riflagan va anatomiyasi anchagina toʻliqroq holatda Thomas Willis tomonidan 1664-yilda yoritilgan. 18-asr davomida anatomiya boʻyicha koʻp ishlar qilingan, lekin 19-asr boshlaridagina miyacha funksiyasi boʻyicha ilk ma’lumotlar olindi. Luigi Rolando 1809-yilda miyacha zararlanishi harakatdagi buzilishlarga sabab boʻlishi haqidagi muhim topilmani ma’lum qildi. Jean Pierre Flourens 19-asrning birinchi yarmida batafsil tajriba ishlarini olib bordi. Bu tajribalar miyacha zararlanishiga ega hayvonlar hamon harakatlana olishi mumkinligi, lekin koordinatsiya yoʻqolishi (gʻalati harakatlar, qoʻpol qadamlar va muskullar kuchsizlanishi)ni, shuningdek, zararlanish juda katta boʻlmasa deyarli tiklanishi mumkinligi koʻrsatdi.[87] 20-asr boshlanganida, miyachaning asosiy funksiyasi harakat boshqariluvi bilan bogʻliqligi koʻpchilik tomonidan tasdiqlangandi; 20-asr birinchi yarmida odamdagi miyacha kasalliklari klinik simptomlarining bir qancha batafsil tavsiflari yaratildi.[4]
Manbalar
[tahrir | manbasini tahrirlash]- ↑ Hodos, William (2009), „Evolution of Cerebellum“, Encyclopedia of Neuroscience, Springer, 1240–1243-bet, doi:10.1007/978-3-540-29678-2_3124, ISBN 978-3-540-23735-8
- ↑ Wolf U, Rapoport MJ, Schweizer TA (2009). „Evaluating the affective component of the cerebellar cognitive affective syndrome“. Journal of Neuropsychiatry and Clinical Neurosciences. 21-jild, № 3. 245–53-bet. doi:10.1176/jnp.2009.21.3.245. PMID 19776302.
- ↑ Schmahmann, Jeremy D.; Caplan, David (1-fevral 2006-yil). „Cognition, emotion and the cerebellum“. Brain (inglizcha). 129-jild, № 2. 290–292-bet. doi:10.1093/brain/awh729. ISSN 0006-8950. PMID 16434422.
{{cite magazine}}: CS1 maint: date format () - ↑ 4,0 4,1 4,2 4,3 Fine EJ, Ionita CC, Lohr L (December 2002). „The history of the development of the cerebellar examination“. Seminars in Neurology. 22-jild, № 4. 375–84-bet. doi:10.1055/s-2002-36759. PMID 12539058.
- ↑ 5,0 5,1 Purves, Dale. Neuroscience, 5th, Sunderland, Mass.: Sinauer, 2011 — 417–423-bet. ISBN 978-0-87893-695-3.
- ↑ 6,0 6,1 6,2 Albus JS (1971). „A theory of cerebellar function“. Math. Biosciences. 10-jild, № 1–2. 25–61-bet. CiteSeerX 10.1.1.14.7524. doi:10.1016/0025-5564(71)90051-4.
- ↑ Purves, Dale Purves. Neuroscience., 4th, New York: W. H. Freeman, 2007 — 197–200-bet. ISBN 978-0-87893-697-7.
- ↑ „Chapter 20“, Gray's anatomy : the anatomical basis of clinical practice, 40th Susan Standring: , London: Churchill Livingstone, 2008 — 297-bet. ISBN 978-0-8089-2371-8.
- ↑ 9,00 9,01 9,02 9,03 9,04 9,05 9,06 9,07 9,08 9,09 9,10 9,11 9,12 9,13 9,14 9,15 9,16 9,17 9,18 9,19 9,20 9,21 9,22 9,23 9,24 „Ch. 7 Cerebellum“, The Synaptic Organization of the Brain Shepherd GM: . New York: Oxford University Press, 2004. ISBN 978-0-19-515955-4.
- ↑ Herculano-Houzel S (2010). „Coordinated scaling of cortical and cerebellar numbers of neurons“. Frontiers in Neuroanatomy. 4-jild. 12-bet. doi:10.3389/fnana.2010.00012. PMC 2839851. PMID 20300467.
- ↑ 11,00 11,01 11,02 11,03 11,04 11,05 11,06 11,07 11,08 11,09 „The cerebellum“, Principles of Neural Science, 2nd edition. New York: Elsevier, 1985 — 502–522-bet.
- ↑ Snider, Raymond; Stowell, Averill (1-noyabr 1944-yil). „Receiving Areas of the Tactile, Auditory, and Visual Systems in the Cerebellum“. Journal of Neurophysiology. 7-jild, № 6. 331–357-bet. doi:10.1152/jn.1944.7.6.331.
{{cite magazine}}: CS1 maint: date format () - ↑ Concise Text of Neuroscience, 2nd, Lippincott Williams & Wilkins, 2000. ISBN 978-0-683-30460-2.
- ↑ Petersen SE, Fox PT, Posner MI, Mintun M, Raichle ME (1989). „Positron emission tomographic studies of the processing of single words“. Journal of Cognitive Neuroscience. 1-jild, № 2. 153–70-bet. doi:10.1162/jocn.1989.1.2.153. PMID 23968463.
- ↑ Timmann D, Daum I (2007). „Cerebellar contributions to cognitive functions: a progress report after two decades of research“. Cerebellum. 6-jild, № 3. 159–62-bet. doi:10.1080/14734220701496448. PMID 17786810.
- ↑ Strick PL, Dum RP, Fiez JA (2009). „Cerebellum and nonmotor function“. Annual Review of Neuroscience. 32-jild. 413–34-bet. doi:10.1146/annurev.neuro.31.060407.125606. PMID 19555291.
- ↑ Buckner RL (October 2013). „The cerebellum and cognitive function: 25 years of insight from anatomy and neuroimaging“. Neuron. 80-jild, № 3. 807–15-bet. doi:10.1016/j.neuron.2013.10.044. PMID 24183029.
- ↑ Schweighofer N, Doya K, Kuroda S (March 2004). „Cerebellar aminergic neuromodulation: towards a functional understanding“. Brain Research. Brain Research Reviews. 44-jild, № 2–3. 103–16-bet. doi:10.1016/j.brainresrev.2003.10.004. PMID 15003388.
- ↑ Felizola SJ, Nakamura Y, Ono Y, Kitamura K, Kikuchi K, Onodera Y, Ise K, Takase K, Sugawara A, Hattangady N, Rainey WE, Satoh F, Sasano H (April 2014). „PCP4: a regulator of aldosterone synthesis in human adrenocortical tissues“. Journal of Molecular Endocrinology. 52-jild, № 2. 159–67-bet. doi:10.1530/JME-13-0248. PMC 4103644. PMID 24403568.
- ↑ Eccles JC, Llinás R, Sasaki K (January 1966). „The excitatory synaptic action of climbing fibres on the Purkinje cells of the cerebellum“. Journal of Physiology. 182-jild, № 2. 268–96-bet. doi:10.1113/jphysiol.1966.sp007824. PMC 1357472. PMID 5944665.
- ↑ 21,0 21,1 21,2 21,3 Simpson JI, Wylie DR, De Zeeuw CI (1996). „On climbing fiber signals and their consequence(s)“. Behav. Brain Sci. 19-jild, № 3. 384–398-bet. doi:10.1017/S0140525X00081486.
- ↑ Whitney ER, Kemper TL, Rosene DL, Bauman ML, Blatt GJ (February 2008). „Calbindin-D28k is a more reliable marker of human Purkinje cells than standard Nissl stains: a stereological experiment“. Journal of Neuroscience Methods. 168-jild, № 1. 42–7-bet. doi:10.1016/j.jneumeth.2007.09.009. PMID 17961663.
- ↑ 23,0 23,1 Rusanescu G, Mao J (February 2017). „Peripheral nerve injury induces adult brain neurogenesis and remodelling“. Journal of Cellular and Molecular Medicine. 21-jild, № 2. 299–314-bet. doi:10.1111/jcmm.12965. PMC 5264155. PMID 27665307.
- ↑ 24,0 24,1 24,2 Marr D (June 1969). „A theory of cerebellar cortex“. Journal of Physiology. 202-jild, № 2. 437–70-bet. doi:10.1113/jphysiol.1969.sp008820. PMC 1351491. PMID 5784296.
- ↑ 25,00 25,01 25,02 25,03 25,04 25,05 25,06 25,07 25,08 25,09 25,10 Apps R, Garwicz M (April 2005). „Anatomical and physiological foundations of cerebellar information processing“. Nature Reviews. Neuroscience. 6-jild, № 4. 297–311-bet. doi:10.1038/nrn1646. PMID 15803161.
- ↑ Manni E, Petrosini L (March 2004). „A century of cerebellar somatotopy: a debated representation“. Nature Reviews. Neuroscience. 5-jild, № 3. 241–9-bet. doi:10.1038/nrn1347. PMID 14976523.
- ↑ Oscarsson O (1979). „Functional units of the cerebellum-sagittal zones and microzones“. Trends Neurosci. 2-jild. 143–145-bet. doi:10.1016/0166-2236(79)90057-2.
- ↑ The Handbook of Cognitive Neuropsychology: What Deficits Reveal about the Human Mind. Psychology Press, 2001 — 481-bet. ISBN 978-1-84169-044-5.
- ↑ 29,0 29,1 Doya K (December 2000). „Complementary roles of basal ganglia and cerebellum in learning and motor control“. Current Opinion in Neurobiology. 10-jild, № 6. 732–9-bet. doi:10.1016/S0959-4388(00)00153-7. PMID 11240282.
- ↑ Manto M, Mariën P (2015). „Schmahmann's syndrome - identification of the third cornerstone of clinical ataxiology“. Cerebellum & Ataxias. 2-jild. 2-bet. doi:10.1186/s40673-015-0023-1. PMC 4552302. PMID 26331045.
- ↑ Schmahmann JD, Sherman JC (April 1998). „The cerebellar cognitive affective syndrome“. Brain. 121-jild, № 4. 561–79-bet. doi:10.1093/brain/121.4.561. PMID 9577385.
- ↑ Levisohn L, Cronin-Golomb A, Schmahmann JD (May 2000). „Neuropsychological consequences of cerebellar tumour resection in children: cerebellar cognitive affective syndrome in a paediatric population“. Brain. 123-jild, № 5. 1041–50-bet. doi:10.1093/brain/123.5.1041. PMID 10775548.
- ↑ Buckner RL, Krienen FM, Castellanos A, Diaz JC, Yeo BT (November 2011). „The organization of the human cerebellum estimated by intrinsic functional connectivity“. Journal of Neurophysiology. 106-jild, № 5. 2322–45-bet. doi:10.1152/jn.00339.2011. PMC 3214121. PMID 21795627.
- ↑ Hernáez-Goñi P, Tirapu-Ustárroz J, Iglesias-Fernández L, Luna-Lario P (November 2010). „Participación del cerebelo en la regulación del afecto, la emoción y la conducta“ [The role of the cerebellum in the regulation of affection, emotion and behavior]. Revista de Neurología (Spanish). 51-jild, № 10. 597–609-bet. doi:10.33588/rn.5110.2010394. PMID 21069639.
{{cite magazine}}: CS1 maint: unrecognized language () - ↑ Turner BM, Paradiso S, Marvel CL, Pierson R, Boles Ponto LL, Hichwa RD, Robinson RG (March 2007). „The cerebellum and emotional experience“. Neuropsychologia. 45-jild, № 6. 1331–41-bet. doi:10.1016/j.neuropsychologia.2006.09.023. PMC 1868674. PMID 17123557.
- ↑ Doya K (October 1999). „What are the computations of the cerebellum, the basal ganglia and the cerebral cortex?“. Neural Networks. 12-jild, № 7–8. 961–974-bet. doi:10.1016/S0893-6080(99)00046-5. PMID 12662639.
- ↑ The Cerebellum as a Neuronal Machine. Springer-Verlag, 1967.
- ↑ The Cerebellum as a Neuronal Machine, p. 311
- ↑ 39,0 39,1 Boyden ES, Katoh A, Raymond JL (2004). „Cerebellum-dependent learning: the role of multiple plasticity mechanisms“. Annual Review of Neuroscience. 27-jild. 581–609-bet. doi:10.1146/annurev.neuro.27.070203.144238. PMID 15217344.
- ↑ 40,0 40,1 Houk JC, Buckingham JT, Barto AG (1996). „Models of the cerebellum and motor learning“ (PDF). Behav. Brain Sci. 19-jild, № 3. 368–383-bet. CiteSeerX 10.1.1.118.2997. doi:10.1017/S0140525X00081474. 2017-08-09da asl nusxadan (PDF) arxivlandi.
- ↑ Fujita M (1982). „Adaptive filter model of the cerebellum“. Biological Cybernetics. 45-jild, № 3. 195–206-bet. doi:10.1007/BF00336192. PMID 7171642.
- ↑ Gilbert PF, Thach WT (June 1977). „Purkinje cell activity during motor learning“. Brain Research. 128-jild, № 2. 309–28-bet. doi:10.1016/0006-8993(77)90997-0. PMID 194656.
- ↑ Christian KM, Thompson RF (2003). „Neural substrates of eyeblink conditioning: acquisition and retention“. Learning & Memory. 10-jild, № 6. 427–55-bet. doi:10.1101/lm.59603. PMID 14657256.
- ↑ Braitenberg V, Atwood RP (February 1958). „Morphological observations on the cerebellar cortex“. Journal of Comparative Neurology. 109-jild, № 1. 1–33-bet. doi:10.1002/cne.901090102. PMID 13563670.
- ↑ Braitenberg V, Heck D, Sultan F (June 1997). „The detection and generation of sequences as a key to cerebellar function: experiments and theory“. Behavioral and Brain Sciences. 20-jild, № 2. 229–45, discussion 245–77-bet. doi:10.1017/s0140525x9700143x. PMID 10096998.
- ↑ Ivry RB, Spencer RM, Zelaznik HN, Diedrichsen J (December 2002). „The cerebellum and event timing“. Annals of the New York Academy of Sciences. 978-jild, № 1. 302–17-bet. Bibcode:2002NYASA.978..302I. doi:10.1111/j.1749-6632.2002.tb07576.x. PMID 12582062.
- ↑ Pellionisz A, Llinás R (1982). „Space-time representation in the brain. The cerebellum as a predictive space-time metric tensor“. Neuroscience. 7-jild, № 12. 2949–70-bet. doi:10.1016/0306-4522(82)90224-X. PMID 7162624.
- ↑ Horváth G (2003). „CMAC: Reconsidering an old neural network“ (PDF). Intelligent Control Systems and Signal Processing. 2020-05-20da asl nusxadan (PDF) arxivlandi. Qaraldi: 2009-12-24.
- ↑ „Examination of Coordination“, Physical Rehabilitation. Philadelphia: F. A. Davis, 2007 — 193–225-bet.
- ↑ Mariën P, Manto M. The linguistic cerebellum. London, UK: Academic Press, 2016 — 337–351-bet. ISBN 978-0-12-801608-4.
- ↑ Gilman S (March 1998). „Imaging the brain. Second of two parts“. New England Journal of Medicine. 338-jild, № 13. 889–96-bet. doi:10.1056/NEJM199803263381307. PMID 9516225.
- ↑ 52,0 52,1 „NINDS Ataxias and Cerebellar or Spinocerebellar Degeneration Information Page“. National Institutes of Health (16-aprel 2014-yil). 9-fevral 2015-yilda asl nusxadan arxivlangan. Qaraldi: 2-fevral 2015-yil.(Wayback Machine saytida 2015-02-09 sanasida arxivlangan)
- ↑ „Veterans of Iraq, Afghanistan Show Brain Changes Related to Explosion Exposure“. Scientific American (15-yanvar 2016-yil). 20-yanvar 2016-yilda asl nusxadan arxivlangan. Qaraldi: 21-yanvar 2016-yil.
- ↑ Vincent M, Hadjikhani N (June 2007). „The cerebellum and migraine“. Headache. 47-jild, № 6. 820–33-bet. doi:10.1111/j.1526-4610.2006.00715.x. PMC 3761082. PMID 17578530.
- ↑ 55,0 55,1 „NINDS Cerebellar Degeneration Information Page“. National Institutes of Health (28-fevral 2014-yil). 18-fevral 2015-yilda asl nusxadan arxivlangan. Qaraldi: 2-fevral 2015-yil.(Wayback Machine saytida 2016-12-02 sanasida arxivlangan)
- ↑ Horvath S, Mah V, Lu AT, Woo JS, Choi OW, Jasinska AJ, Riancho JA, Tung S, Coles NS, Braun J, Vinters HV, Coles LS (May 2015). „The cerebellum ages slowly according to the epigenetic clock“ (PDF). Aging. 7-jild, № 5. 294–306-bet. doi:10.18632/aging.100742. PMC 4468311. PMID 26000617. 2015-05-25da asl nusxadan arxivlandi (PDF). (Wayback Machine saytida 2016-03-18 sanasida arxivlangan)
- ↑ Fraser HB, Khaitovich P, Plotkin JB, Pääbo S, Eisen MB (September 2005). „Aging and gene expression in the primate brain“. PLOS Biology. 3-jild, № 9. e274-bet. doi:10.1371/journal.pbio.0030274. PMC 1181540. PMID 16048372.
- ↑ Andersen BB, Gundersen HJ, Pakkenberg B (November 2003). „Aging of the human cerebellum: a stereological study“. Journal of Comparative Neurology. 466-jild, № 3. 356–65-bet. doi:10.1002/cne.10884. PMID 14556293.
- ↑ Raz N, Gunning-Dixon F, Head D, Williamson A, Acker JD (2001). „Age and sex differences in the cerebellum and the ventral pons: a prospective MR study of healthy adults“ (PDF). American Journal of Neuroradiology. 22-jild, № 6. 1161–7-bet. PMID 11415913. 2008-12-17da asl nusxadan arxivlandi (PDF).
- ↑ 60,0 60,1 60,2 60,3 The Merck Manual of Diagnosis and Therapy, 18th, Whitehouse Station, New Jersey: Merck Research Libraries, 2006 — 1886–1887-bet.
- ↑ Campbell J, Gilbert WM, Nicolaides KH, Campbell S (August 1987). „Ultrasound screening for spina bifida: cranial and cerebellar signs in a high-risk population“. Obstetrics and Gynecology. 70-jild, № 2. 247–50-bet. PMID 3299184.
- ↑ 62,0 62,1 Hatten ME, Heintz N (1995). „Mechanisms of neural patterning and specification in the developing cerebellum“. Annual Review of Neuroscience. 18-jild. 385–408-bet. doi:10.1146/annurev.ne.18.030195.002125. PMID 7605067.
- ↑ Polkinghorn WR, Tarbell NJ (May 2007). „Medulloblastoma: tumorigenesis, current clinical paradigm, and efforts to improve risk stratification“. Nature Clinical Practice. Oncology. 4-jild, № 5. 295–304-bet. doi:10.1038/ncponc0794. PMID 17464337.
- ↑ Cerebellum development and medulloblastoma, 2011 — 235–82-bet. DOI:10.1016/B978-0-12-380916-2.00008-5. ISBN 9780123809162.
- ↑ „NINDS Joubert Syndrome Information Page“. National Institutes of Health (23-dekabr 2013-yil). 4-yanvar 2015-yilda asl nusxadan arxivlangan. Qaraldi: 9-yanvar 2015-yil.(Wayback Machine saytida 2016-12-08 sanasida arxivlangan)
- ↑ „NINDS Dandy-Walker Information Page“. National Institutes of Health (14-fevral 2014-yil). 4-yanvar 2015-yilda asl nusxadan arxivlangan. Qaraldi: 9-yanvar 2015-yil.(Wayback Machine saytida 2016-12-02 sanasida arxivlangan)
- ↑ „NINDS Cerebellar Hypoplasia Information Page“. National Institutes of Health (29-sentabr 2011-yil). 4-yanvar 2015-yilda asl nusxadan arxivlangan. Qaraldi: 9-yanvar 2015-yil.(Wayback Machine saytida 2016-12-09 sanasida arxivlangan)
- ↑ „Chiari Malformation Fact Sheet“. National Institutes of Health (10-dekabr 2014-yil). 27-oktabr 2011-yilda asl nusxadan arxivlangan. Qaraldi: 9-yanvar 2015-yil. (Wayback Machine saytida 2011-10-27 sanasida arxivlangan)
- ↑ „NINDS Dyssynergia Cerebellaris Myoclonica Information Page“. National Institutes of Health (14-fevral 2011-yil). 16-fevral 2015-yilda asl nusxadan arxivlangan. Qaraldi: 1-fevral 2015-yil.(Wayback Machine saytida 2015-02-16 sanasida arxivlangan)
- ↑ „NINDS Olivopontocerebellar Atrophy Information Page“. National Institutes of Health (16-aprel 2014-yil). 27-yanvar 2012-yilda asl nusxadan arxivlangan. Qaraldi: 9-yanvar 2015-yil.(Wayback Machine saytida 2012-01-27 sanasida arxivlangan)
- ↑ „NINDS Paraneoplastic Syndromes Information Page“. National Institutes of Health (12-mart 2009-yil). 4-yanvar 2015-yilda asl nusxadan arxivlangan. Qaraldi: 9-yanvar 2015-yil.(Wayback Machine saytida 2015-01-04 sanasida arxivlangan)
- ↑ „NINDS Wernicke-Korsakoff Syndrome Information Page“. National Institutes of Health (14-fevral 2007-yil). 4-yanvar 2015-yilda asl nusxadan arxivlangan. Qaraldi: 9-yanvar 2015-yil.(Wayback Machine saytida 2016-12-12 sanasida arxivlangan)
- ↑ Moulton EA, Schmahmann JD, Becerra L, Borsook D (October 2010). „The cerebellum and pain: passive integrator or active participator?“. Brain Research Reviews. 65-jild, № 1. 14–27-bet. doi:10.1016/j.brainresrev.2010.05.005. PMC 2943015. PMID 20553761.
- ↑ Baumann O, Borra RJ, Bower JM, Cullen KE, Habas C, Ivry RB, Leggio M, Mattingley JB, Molinari M, Moulton EA, Paulin MG, Pavlova MA, Schmahmann JD, Sokolov AA (April 2015). „Consensus paper: the role of the cerebellum in perceptual processes“. Cerebellum. 14-jild, № 2. 197–220-bet. doi:10.1007/s12311-014-0627-7. PMC 4346664. PMID 25479821.
- ↑ 75,0 75,1 75,2 75,3 Bell CC, Han V, Sawtell NB (2008). „Cerebellum-like structures and their implications for cerebellar function“. Annual Review of Neuroscience. 31-jild. 1–24-bet. doi:10.1146/annurev.neuro.30.051606.094225. PMID 18275284.
- ↑ Woodhams PL (July 1977). „The ultrastructure of a cerebellar analogue in octopus“. Journal of Comparative Neurology. 174-jild, № 2. 329–45-bet. doi:10.1002/cne.901740209. PMID 864041.
- ↑ 77,0 77,1 The Vertebrate Body. Philadelphia: Holt-Saunders International, 1977 — 531-bet. ISBN 978-0-03-910284-5.
- ↑ Shi Z, Zhang Y, Meek J, Qiao J, Han VZ (August 2008). „The neuronal organization of a unique cerebellar specialization: the valvula cerebelli of a mormyrid fish“. Journal of Comparative Neurology. 509-jild, № 5. 449–73-bet. doi:10.1002/cne.21735. PMC 5884697. PMID 18537139.
- ↑ Weaver AH (March 2005). „Reciprocal evolution of the cerebellum and neocortex in fossil humans“. Proceedings of the National Academy of Sciences of the United States of America. 102-jild, № 10. 3576–80-bet. Bibcode:2005PNAS..102.3576W. doi:10.1073/pnas.0500692102. PMC 553338. PMID 15731345.
- ↑ 80,0 80,1 80,2 Schoenemann PT (1-dekabr 2009-yil). „Evolution of Brain and Language“. Language Learning. 59-jild. 162–186-bet. doi:10.1111/j.1467-9922.2009.00539.x.
{{cite magazine}}: CS1 maint: date format () - ↑ MacLeod CE, Zilles K, Schleicher A, Rilling JK, Gibson KR (April 2003). „Expansion of the neocerebellum in Hominoidea“. Journal of Human Evolution. 44-jild, № 4. 401–29-bet. doi:10.1016/S0047-2484(03)00028-9. PMID 12727461.
- ↑ Roberts PD, Portfors CV (June 2008). „Design principles of sensory processing in cerebellum-like structures. Early stage processing of electrosensory and auditory objects“. Biological Cybernetics. 98-jild, № 6. 491–507-bet. doi:10.1007/s00422-008-0217-1. PMID 18491162.
- ↑ Is the cerebellum sensory for motor's sake, or motor for sensory's sake: the view from the whiskers of a rat?, 1997 — 463–96-bet. DOI:10.1016/S0079-6123(08)63381-6. ISBN 978-0-444-82313-7.
- ↑ Heiney SA, Kim J, Augustine GJ, Medina JF (February 2014). „Precise control of movement kinematics by optogenetic inhibition of Purkinje cell activity“. Journal of Neuroscience. 34-jild, № 6. 2321–30-bet. doi:10.1523/JNEUROSCI.4547-13.2014. PMC 3913874. PMID 24501371.
- ↑ Witter L, Canto CB, Hoogland TM, de Gruijl JR, De Zeeuw CI (2013). „Strength and timing of motor responses mediated by rebound firing in the cerebellar nuclei after Purkinje cell activation“. Frontiers in Neural Circuits. 7-jild. 133-bet. doi:10.3389/fncir.2013.00133. PMC 3748751. PMID 23970855.
- ↑ „Ch. 11: Cerebellum“, The Human Brain and Spinal Cord, 2nd, Norman Publishing, 1996 — 629-bet. ISBN 978-0-930405-25-0.
- ↑ Ito M (December 2002). „Historical review of the significance of the cerebellum and the role of Purkinje cells in motor learning“. Annals of the New York Academy of Sciences. 978-jild, № 1. 273–88-bet. Bibcode:2002NYASA.978..273I. doi:10.1111/j.1749-6632.2002.tb07574.x. PMID 12582060.
Havolalar
[tahrir | manbasini tahrirlash]| Vikiomborda Miyacha haqida turkum mavjud |
- Cerebellum Rodolfo Llinas and Mario N. Negrello, Scholarpedia, 10(1):4606.
- Cerebellum–Cell Centered Database
- Handbook of the Cerebellum and Cerebellar Disorders – Manto, M., Gruol, D.L., Schmahmann, J., Koibuchi, N., Rossi, F. (Eds.) – Springer – New York
- Andoza:BrainMaps
- A Man's Incomplete Brain Reveals Cerebellum's Role In Thought And Emotion
- Woman living without cerebellum
- Essentials of Cerebellum and Cerebellar Disorders. Gruol, D.L., Koibuchi, N., Manto, M., Molinari, M., Schmahmann, J.D., Shen, Y. (Eds.). Springer, New York, 2016
- Cerebellum histology images
- The Cerebellum – Journal (Springer Nature)
- Cerebellum and Ataxias – Journal (BioMed Central)