Krvna grupa

Prvi red: krvne grupe
Drugi red: antitijela u plazmi
Treći red: antigeni na eritrocitima
Krvna grupa je jedna od klasifikacijskih odrednica krvi, a bazirana je na prisutnosti ili nedostatku naslijeđene antigenske supstance na površini eritrocita. Ovi antigeni mogu biti proteini, ugljikohidrati, glikoproteini ili glikolipidi, u zavisnosti od sistema krvnih grupa, a neki od ovih antigena su prisutni i na površinama drugih vrsta ćelija raznih tkiva.[1]
Suglasno specifičnosti antigena i/ili antitijela u krvi, fenotipovi krvnih grupa se klasificiraju po više sistema. Neki su toliko rijetki da se označavaju kao privatni (engleski: private), dok su drugi javni (engleski: public), manje ili više rasprostranjeni u većem dijelu svjetske populacije.[2]
Sistemi krvnih grupa
[uredi | uredi izvor]Opća imunogenetička svojstva krvnih antigena
[uredi | uredi izvor]Sistemi krvnih grupa pripadaju skupini kvalitativnih biohemijsko–fizioloških – monogenskih svojstava (osobina), čiji su genetički determinatori locirani na autosomima (autosomni) ili heterosomima (heterosomni, gonosomni). U ovoj skupini – imunogenetički značajnih sistema antigena – javljaju se tri kategorije nasljeđivanja: ABO sistem krvnih grupa kontrolira serija od tri glavna (multipla) alela, a individualnu pripadnost jednom od mogućih fenotipova Rh sistema određuju aleli tri blisko vezana genska lokusa humane hromosomske garniture. Istovremeno, krvne grupe MN sistema određuje jedan genski lokus među čijim alelima nema odnosa dominantnosti, tj. karakterizira ih kodominantnost. Pojava svakog od tih alela u genotipu testirane osobe se manifestira obligatnim prisustvom antigena koji je pod njihovom kontrolom. Za razliku od ovih sistema nasljedne varijacije, prema općeprihvaćenim teorijama, mnoge morfološke monogenske osobine se tek konvencionalno mogu svrstati u tu kategoriju.[3] [4]
U ostalim sistemima krvnih grupa moguće varijante (krvne grupe) se nasljeđuju po jednom od pomenutih modela ili se javljaju u mnoštvu izoantigena.
U ukupnom svjetskom stanovništvu, imunogenetička istraživanja su registrirala petnaestak sistema širokorasprostranjenih (eng. public) sistema krvnih grupa, sa respektabilnom frekvencijom mogućih alternativnih fenotipova. Pored toga, sporadično – kao svojevrsni rariteti – u ograničenom broju individua i njihovih porodica – javljaju se i krvni antigeni osobenih (eng. private) sistema grupne specifičnosti.
Interpretacija prirode interakcije među genetičkim determinatorima krvnih grupa različitih sistema je relativno jednostavna. U nauci o biološkom nasljeđivanju ne postoji oblast sa tako jasnim odrednicama kao što je to slučaj sa genetikom humanih krvnih grupa. Iako su neki njeni segmenti doista složeni, oni se više tiču serologije nego genetike.
- Individualna pripadnost krvnim grupama, tj. različiti antigeni koji se nalaze na površini eritrocita, genetički su određeni. Praktično i nema sredinskih uticaja na njihovo ispoljavanje koji bi zakomplicirali mogućnost pune genske ekspresije, a nekoliko nedavno potvrđenih izuzetaka su ekstremno rijetki fenomeni.
- Konkordantnost između prisutnog alelogena u genotipu i antigena na eritrocitima je gotovo uvijek potpuna (100%). Reakcija sa specifičnim antitijelima redovno dokazuje prisustvo odgovarajućeg antigena, što istovremeno dokazuje i prisustvo kodirajućeg gena. Pritom reakcija sa komplementarnim antiserumom indicira prisustvo tog gena, bez obzira na eventualno prisustvo drugih.
- Interakcija i ekspresija alelnih gena za mnoge druge osobine može se zakomplicirati do stupnja na kojem posmatrani fenotipski sistem postaje nepodesan za genetičku analizu. Kada su u pitanju geni koji određuju krvne grupe, ovakvih interakcija je veoma malo, a kada se i pojave, prisustvo prisutnih gena se gotovo uvijek može registrirati.
- U nasljeđivanju krvnih grupa nema komplikacija uzrokovanih drugim genima koji imaju nerazlučivo slične efekte, kao što je to čest slučaj sa nasljeđivanjem kongenitalnih anomalija, npr. Prisustvo antigena A ili B, naprimjer, također je uvijek rezultat aktivnosti istih alelogena, jer ne postoji neki drugi gen čiji bi produkt također reagovao sa anti–A ili anti–B.
- Generalno gledajući, među genetičkim determinatorima krvnih grupa poznatih sistema nema odnosa dominantnosti – recesivnosti. Dosadašnja iskustva pokazuju da se obično otkrije alelomorfni par koji određuje prisustvo dva različita antigena na eritrocitima, nakon čega se, relativno brzo, detektiraju i odgovarajući antiserimi koji sa njima reaguju. Eritrociti svakog homozigota isključivo reagiraju (aglutiniraju) samo sa jednim – specifičnim antiserumom (ali ne i sa drugima!). Istovremeno, eritrociti heterozigota (genotipa sa dva različita alela), reagiraju sa oba antiseruma. Prema tome, ako su nam dostupna oba antiseruma, serološkim testovima, može se pouzdano odrediti genetička konstitucija svake individue.
- Respektirajući sve pomenute činjenice, veoma je bitno permanentno prisustvo svijesti da očita jednostavnost naljeđivanja krvnih grupa ponekad može i zavarati – unijeti konfuziju i obmane, kako u istraživačkoj tako i u dijagnostičkoj praksi. Takve komplikacije se javljaju sporadično, ali i kontinuirano, a sa mogućim teškim posljedicama po zdravlje pacijenata, u transfuzijskim i transplantacijskim tretmanima.
Klasifikacija krvnih grupa
[uredi | uredi izvor]| Sistemi krvnih grupa – priroda epitopa i hromosomska pozicija odgovarajućih gena[5] | |||||
|---|---|---|---|---|---|
| N° | Inicijalni simbol | Nomenklatura
ISBT / HGNC |
Epitop | Hromosomska lokacija | OMIM[6][7] |
| 001 | ABO | ABO | Ugljikohidrat (N-acetilgalaktozamin, galaktoza); A, B i H antigeni uglavnom izazivaju reakcije IgM antitijela,anti-H je vrlo rijetka (pogledajte Hh sistem antigena – Bombay fenotip) | 9q34.2* | OMIM: 110300 |
| 002 | MNS | MNS / GYPA - GYPB - (GYPE) | Glikoforin GPA / GPB (glikoforini A i B), mogući receptori Plamodium falciparum, glavni antigeni: M, N, S, s
Antigeni M i N na GPA (mutacije M→N : ser1leu i gly5glu), S i s lGPB (mutacije S → S: met29thr); u prva 26 N-terminalna AA, ekstramembranozne FAM-N i GPB su identične; neizvjesno postojanje MCP; nedostatak uzrokuje GPA rijetki fenotip En(a) - deficit sijalinske kiseline, a pad u zeta potencijalu. |
4q31.21 | OMIM: 111300 |
| 003 | P | P1PK / A4GALT | Glikolipid; Antigen P1, α1,4galaktoza na paraglobozidu; antigen P1: Galα1-4Galβ1-4GlNacβ1-3Lactozilceramid | 22q13.2 | OMIM: 111400 |
| 004 | Rh | RH / RHD - RHCE | Proteini RHD / RHCE; C, c, D, E, e antigeni (ne postoji "d" antigen - malo slovo d označava odsustvo D proteina, tj. antigena) | 1p36.11 | OMIM: 111680 OMIM: 111700 |
| 005 | Lutheran | LU / BCAM | Protein IgSF (član imunoglobulin superfamilije); set od 21 antigena | 19q13.32 | OMIM: 111200 |
| 006 | Kell | KEL / KEL | Glikoprotein, endopeptidaza; priključna (Cys72) disulfid veza za protein Kx (Cys347); član imunoglobulinska superfamilije); set od 21 antigena | 7q34 | OMIM: 110900 |
| 007 | Lewis | LE / FUT3 | Ugljikohidrat (fukoza, rezidua); glavni antigeni Lea i Leb - asocirani sa sekrecijom tkivnih ABH antigena | 19p13.3 | OMIM: 111100 |
| 008 | Duffy | FY / DARC | Protein (ECR himokin receptor); glavni antigeni: Fya and Fyb. Osobe kojima nedostaje Duffy antigen sasvim su imune na malariju uzrokovanu plazmodijima (Plasmodium vivax i Plasmodium knowlesi) | 1q23.2 | OMIM: 110700 |
| 009 | Kidd | JK / SLC14A1 | Protein (urea transporter); glavni antigeni: Jka i Jkb | 18q12.3 | OMIM: 111000 |
| 010 | Diego | DI / SLC4A1 | Glikoprotein (band 3, AE 1, ili anion razmjene); pozitivna krv nađena samo u Istočnoj Aziji i kod autohtonih sjevernoameričkih Indijanaca. | 17q21.31* | OMIM: 110500 |
| 011 | Yt sistem antigena/Cartwright | YT / ACHE | Protein (AChE, acetilholinesteraza), fiksiran na membrane ili glikozo-fosfatidilinostitol (GPI) Kell sistema: lod skor = 3.48 za Θ = 0.28 | 7q22.1 | OMIM: 112100 |
| 012 | Xg | XG / XG | Glikoprotein | Xp22.3 | OMIM: 314700 |
| 013 | Scianna | SC / ERMAP | Glikoprotein | 1p34.2 | OMIM: 111750 |
| 014 | Dombrock | DO / ART4 | Glikoprotein ADP-ribotransferaza (fiksiran na ćelijskoj membrani putem GPI ili glicozil-fosfatidil-inositola) | 12p12.3 | OMIM: 110600 |
| 015 | Colton | CO / AQP1 | Akvaporin 1; glavni antigeni: Co(a) i Co(b) | 7p14.1 | OMIM: 110450 |
| 016 | Landsteiner-Wiener | LW /ICAM4 | Protein IgSF (član imunoglobulinske superfamilije) | 19p13.2 | OMIM: 111250 |
| 017 | Chido/Rodgers | CH/RG / C4A - C4B | Komplement C4A C4B (frakcije); 99 % homologije između dva proteina | 6p21.3 | OMIM: 120810 OMIM: 120820 |
| 018 | Hh | H / FUT1 | Ugljikohidrat (fukoza, ostatak) | 19q13.33 | OMIM: 211100 |
| 019 | Kx | XK / XK | Glikoprotein; povezan je (Cys347) po disulfid mostu na Kell protein (Cys72). | Xp21.1 | OMIM: 314850 |
| 020 | Gerbich | GE / GYPC | Glikoforini C i D (GPC / GPD (glikoforini C i D, GPD su rezultanti GPC za brisanje receptora Plasmodium falciparum | 2q14.3 | OMIM: 110750 |
| 021 | Cromer | CROM / DAF | Glikoprotein (DAF ili CD55), regulira dopunu frakcija C3 i C5, pričvršćeni za membranu GPI | 1q32.2 | OMIM: 125240 |
| 022 | Knops | KN / CR1 | Glikoprotein (CR1 ili CD35), receptor imunog kompleksa | 1q32.2 | OMIM: 607486 |
| 023 | Indian | IN / CD44 | Glikoprotein CD44 adhezijske funkcije (ligand hialuronične kiseline); mutacija IN1 ⇒ IN2: Pro46Arg; genske frekvencije – IN1:0,014; IN2:0,986 | 11p13 | OMIM: 609027 |
| 024 | OK | OK / BSG | Glikoprotein (CD147), član imunoglobulinske natporodice | 19p13.3 | OMIM: 111380 |
| 025 | Raph | MER2 / CD151 | Tetraspanin CD151, transmembranski glikoprotein | 11p15.5 | OMIM: 179620 OMIM: 602243 |
| 026 | JMHG | JMH / SEMA7A | Protein (fiksiran za membranu pomoću GPI) | 15q24.1 | OMIM: 607961 |
| 027 | Ii | I / GCNT2 | Polisaharid (poli-N-acetilactozaminoglikan) – razgranat (račvast) - I / nerazgranat (jednostavan) - i | 6p24.2 | OMIM: 110800 |
| 028 | Globozid | GLOB / B3GALNT1 | Glikolipid, acetil-galactoza-aminil-transferaza 1; antigen P
Antigène [P]: GalNAcβ1-3Galα1]-4Galβ1-4Glβ1-1-keramid receptor parvovirus B19 ; laktozilceramid : Galβ1-4Glβ1-1-keramid, supstrat za P1. |
3q26.1 | OMIM: 603094 |
| 029 | GIL | GIL / AQP3 | Akvaporin 3 | 9p13.3 | OMIM: 607457 |
| 030 | Rh-asocirani glikoprotein | RHAG / RHAG | Glikoprotein, 409aa, 36 % homologije svoje RHCED. Esencijalno za Rh-asocirani glikoproteinsku ekspresiju HR. | 6p12.3 | OMIM: 180297 |
| 031 | Forssman | FORS / GBGT1 | Glikozil-transferaza; transferira α1-3 N-acetil-galaktozamin na globozid(antigen D) | 9q34.2 | OMIM: 606074 |
| 032 | Junior/JR | JR / ABCG2 | Protein, 16 egzona, 655 AA, 72 000 Daltona, 3 lokusa N, moguća glikolizacija (šest membranskih pasaža), ABCG2, Multi-poteinski transporter (lijekova), frekvencija Jra+ >99%. | 4q22.1 | OMIM: 614490 |
| 033 | Langereis/LAN | LAN / ABCB6 | Protein, 19 egzona, 842 AA, 80.000 Daltona, 10 cisteina, 4 lokusa; moguće N-glikozilacija, uloga u sintezi hema; ABCB6,porfirin transporter | 2q36 | OMIM: 111600 |
| Najpoznatiji (public) sistemi krvnih grupa i njihovi najfrekventniji izoantigeni | ||||||
|---|---|---|---|---|---|---|
| Sistem krvnih grupa | Izoantigeni | |||||
| ABO | A1, A2, A3, A4, A5, A0, Az, B, O, H | |||||
| Rh | D, C, c, Cw, Cx, E, e, es (VS), Dw, Du, ‘’C’’u, ‘’E’’u, G, ce, Ces (V), Ce, CE, cE, Et, LW | |||||
| MNSs | M, N, S, s, U, Mg, M1, M2, N1, Mc, Ma, Mv, Mk, Tm, Hu, He, Mia, Vw, (Gr), Mur, Hil, Vr, R2a, Sta, Mta, Cla, Nya, Sul, Sj, S2 | |||||
| P | P1, P2, Pk | |||||
| Lutheran | Lua, Lub | |||||
| Lewis | Lea, Leb, Lec, Led | |||||
| Kell | K, k, Kpa, Kpb, Jsa, Jsb | |||||
| Duffi | Fyb, Fyb | |||||
| Kidd | Jka, Jkb, jk | |||||
| Diego | Diab, Dib | |||||
| Yt | I, i | |||||
| Auberger | Aua | |||||
| Xg | Xga | |||||
ABO sistem krvnih grupa
[uredi | uredi izvor]Antigeni i genetika ABO sistema
[uredi | uredi izvor]Antigeni ABO sistema krvnih grupa otkriveni su na osnovu efekata miješanja krvi različitih osoba, odnosno pojave ili odsustva aglutinacije u mogućim kombinacijama eritrocita i seruma ispitanika. Rezultati prvih istraživanja u ovoj oblasti dokazali su da se u humanim crvenim krvnim zrncima nalaze antigene supstance, a u serumu specifična antitijela, koja u međusobnom kontaktu izazivaju izoaglutinaciju (aglutinacija reakcija antigen – antitijelo) među pripadnicima iste vrste organizama), čime je objašnjena fiziološka priroda ovog višeslojno značajnog sistema individualne varijacije, posebno u transplantaciji u najširem smislu pojma (uključujući i transfuziju).
Nakon tri prvootkrivene grupe (A, B, O) ovog sistema, opisana je ubrzo i četvrta (AB) i pouzdano dokazano da se ovaj sistem antigene varijacije diferencira po prisustvu (ili odsustvu) supstanci A i B (na eritrocitima) i odgovarajućih specifičnih antitijela: anti–A i anti–B (u serumu). Također je konstatirano da su, za razliku od većine ostalih sistema krvnih grupa (Rhi MN, naprimjer), antitijela za ove antigene (izuzimajući grupu AB) redovno prisutna u normalnom serumu. Oznake krvnih grupa ovog sistema, po zvaničnoj i danas općeprihvaćenoj nomenklaturi (koja se ranije bazirala i na slijedu rednih brojeva I – IV), indiciraju prisustvo specifičnih antigena na crvenim krvnim zrncima: krvna grupa A sadrži antigenu supstancu A, eritrociti fenotipa B nose antigen B; u krvnoj grupi AB nalaze se obje ove grupno–specifične supstance, a u O (nula) – nijedna od njih. Suglasno tome, u serumu krvne grupe A moguće je prisustvo samo antitijela anti–B i obrnuto – grupa B sadrži samo anti–A supstancu, dok serum AB ne sadrži niti jedno od ovih specifičnih antitijela ABO sistema.
U normalnim okolnostima ne dolazi do autoaglutinacije, ali se izoaglutinacija redovno javlja u inkompatibilnim (heterospecifičnim) kombinacijama miješanja krvi različitih osoba, kada dolazi do transfuzijskog šoka praćenog hemolizom. Prema tome, u transfuzijskim i drugim zahvatima, krv pripadnika grupe O se nesmetano može miješati sa krvlju osoba grupa A i B jer u eritrocitima ne posjeduje ni A ni B antigen (univerzalni davaoci). Osobe krvne grupe AB su univerzalni primaoci (jer u serumu nemaju ni anti–A ni anti–B supstance), dok osobe krvnih grupa A i B, pored homospecifične, mogu primati i transfuziju krvi nulte grupe. Međutim, budući da kod takvih transfuzija postoji izvjestan stupanj rizika, favorizuju su aplikacije krvi homospecifičnih, istovjetnih grupa davatelja i primatelja.
Eritrocitni antigeni su glikolipidne prirode (topivi u alkoholu), a (pored krvi) široko su rasprostranjeni i u ostalim tkivima (osim mozga), ali se nikada ne javljaju u sekretima i ekskretima. Druga skupina antigena istih specifičnosti ima glikoproteinsku strukturu (pa su rastvorljivi u vodi) i susreću se u većini tkiva (osim krvi) i izlučevina (pljuvačka, znoj, suze, mlijeko, želučani sok, gnoj, mokraća, izmet itd.). Na temelju prisistva ili odsustva antigena ABO(H) sistema, osobe se dijele na sekretore i nesekretore. Sposobnost (ne)sekrecije ovih supstanci u te tjelesne izlčevine, genetički je kontrolirana sa nezavisnog genskog lokusa (osobina sekrecija AB(O)H antigena).
Detaljnije analize ovih grupno specifičnih supstanci su pokazale da njihova antigena specifičnost ovisi o strukturi terminalne gliko–komponente odgovarajućih molekula (glikolipida i glikoproteina). U njenoj osnovi je zajednička četveročlana sekvenca prostih šećera, koja također ispoljava antigenu aktivnost (označenu sa H) i koja je prisutna i u fenotipu O. Antigen H ustvari predstavlja prekursornu supstancu u sintezi antigena A i B koji nastaju terminalnim dodavanjem specifičnih prostih aminošećera, dok u krvnoj grpi O on ostaje nealteriran. Samo u izuzetno rijetkim slučajevima, na odgovarajućem lokusu javlja se mutantna varijanta (alel h), koja je izgubila mogućnost kodiranja enzima za katalizu sinteze antigena H iz prekursorne substance. To se manifestira kao Bombay fenotip (svrstan u kategoriju private krvnih grupa). Također treba napomenuti da se u vodi rastvorljivi antigeni ABO(H) sistema u tjelesnim izlučevinama javljaju samo kod "sekretora", dominantne fenotipske oznake u nasljednom dimorfizmu (po) sposobnosti njihove ekskrecije (koji je nezavisan od ABO sistema krvnih grupa).
Antitjela ABO sistema krvnih grupa (anti–A i anti–B) su bjelančevine tipa globulina III–1 frakcije plazme i skupa sa specifičnim antigenima imaju veliki značaj u transfuziji, transplantaciji, forenzičkoj antropologiji i drugim oblastima osnovnih i primijenjenih bioloških nauka.
ABH antigeni, dakle, nisu primarni produkti gena već nastaju u enzimskim reakcijama koje katalizira glikoziltransferaza. Imunodominantne strukture A i B antigena, GanNAc alpha1–>3 (Fuc alpha1–>2) Gal– i Gal alpha1–>3 (Fuc alpha 1–>2) Gal–, nastaju u seriji reakcija; A i B–transferaze koje koiraju funkcionalni aleli (IA i IB aleli) jednog gena ABO lokusa, kataliziraju posljednji korak u sintezi. Protein koji kodira alel IO nije funkcionalan, pa stoga akceptorni supstrat (H antigen: Fuc alpha 1->2 Gal-) ostaje nemodificiran.[8]
ABO alelogeni se sastoje od najmanje sedam egzona, a njihova kodirajuća sekvenca proteže se na 18 kb genomske DNK. Delecija jednog nukleotida u egzonu 6 susreće se u većini (ali ne svim) I0 alela i odgovorna je za gubitak aktivnosti enzima. Prva od sedam supstitucija nukleotida, po kojoj se razlikuju A i B transferaze, nalazi se u egzonu 6. Najveći je egzon 7, koji sadrži preostalih šest supstitucija nukleotida, uzrokuje četiri diferencirajuće aminokiselinske supstitucije transferaza A i B. Među ovim supstitucijama nalaze se i one koje određuju A ili B specifičnost enzima (aminokiselinski ostaci 266 i 268). Pored četiri uobičajena alela (IA1, IA2, IB i I0) identificirani su i brojni aleli koji kodiraju sintezu glikoziltransferaza sa promijenjenom aktivnošću ili specifičnošću.
- ABO lokus je na dugom kraku hromosoma 9: 9q34.
Na osnovu stupnja antigenosti, unutar fenotipa A registrirane su njegove varijante A1, A2, A3, A4 i A5, (sa opadajućim gradijentom antigene aktivnosti), koje se međusobno serološki jasno diferenciraju i ne prepokrivaju. Odnos učestalosti antigena A1, i A2 je oko 4:1; A3, čini manje od 1% učestalosti krvne grupe A; A4 je dvostruko rjeđa, dok je A5 specifičnost pravi raritet.
Osnovu genetičke kontrole prikazane varijacije antigena ABO sistema (prema danas općeprihvaćenoj hipotezi, kojom je oborena prva teorija o dva lokusa) čine tri (multipla) alela (IA, IB, I0) odgovarajućeg genskog lokusa (I), čiju interakciju karakterizira kodominantnost ( IA, IB) u odnosu na recesivni I0, dok u tom paru nema dominantnosti – recesivnosti. Zato se prisustvo svakog od njih u genotipu redovno i fenotipski manifestira pojavom odgovarajućih antigena na eritrocitima. Prema tome, multipli aleli ovog lokusa kombinuju se u šest mogućih genotipova, čiju fenotipsku ekspresiju čine četiri krvne grupe. A i B grupa, dakle, mogu biti (dominantni) homozigoti (IAIA, IBIB) i heterozigoti (IAI0, IB I0); različiti genotipovi iste krvne grupe serološki se ne mogu međusobno diferencirati. Nulta (O) krvna grupa je recesivni homozigot (I0I0), a AB je heterozigotna kombinacija kodominantnih alela (IAIB). Zato (imajući u vidu ovakve relacije genetičkih determinatora ovog sistema antigena), osobe krvnih grupa O i AB u međusobnoj vezi ne mogu imati homospecifično potomstvo (svoje krvne grupe).[9]
Polimorfizam antigena ABO sistema je zabilježen kod mnogih vrsta primata. Tako je uočeno da je kod šimpanzi najfrekventnija krvna grupa A, dok je O mnogo rjeđa, a B krvne grupe uopće nema. Nasuprot tome, gorile imaju samo krvnu grupu B. Na osnovu takvih relacija, pretpostavlja se da determinirajući alelogeni ovih fenotipova kod hominoida odavno nisu pod značajnijim selekcijskim pritiskom. Istovremeno, evolutivno udaljeniji babuni imaju krvne grupe A, B i O, s tom razlikom što su kod čovjeka i ostalih antropomorfnih majmuna ABO(H) antigeni registrirani u crvenim krvnim zrncima, a kod babuna na drugim tipovima ćelija. Iako se i kod njih javlja grupa O, ona je mnogo rjeđa nego kod ljudi. Postoji mogućnost da kod babuna postoji negativna selekcija fenotipa O.
Suglasno prethodnim nalazima, formulirane su dvije teorije evolucione diversifikacije gena za krvne grupe ABO sistema:
- transpecijska evolucija – pretpostavlja da genetička sličnost potiče od zajedničkog primatskog pretka, a
- hipoteza o paralelnoj evoluciji počiva na uvjerenju da su ABO alelogeni istovremeno i nezavisno evoluirali – u paralelnoj filogenezi različitih vrsta primata, u odgovoru na pozitivni selekcijski pritisak.
Druga teorija ima više pristalica zbog prisustva specifičnih mutacija u unutargenskim sekvencama ABO gena.
| Nasljeđivanje genotipova krvnih grupa ABO sistema i njihovo fenotipsko ispoljavanje | |||||||
| Grupa | O | A | B | AB | |||
|---|---|---|---|---|---|---|---|
| Genotip | ii (OO) | IAi (AO) | IAIA (AA) | IBi (BO) | IBIB (BB) | IAIB (AB) | |
| O | ii (OO) | O OO OO OO OO |
O ili A AO OO AO OO |
A AO AO AO AO |
O ili B BO OO BO OO |
B BO BO BO BO |
A ili B AO BO AO BO |
| A | IAi (AO) | O ili A AO AO OO OO |
O ili A AA AO AO OO |
A AA AA AO AO |
O, A, B ili AB AB AO BO OO |
B ili AB AB AB BO BO |
A, B ili AB AA AB AO BO |
| IAIA (AA) | A AO AO AO AO |
A AA AO AA AO |
A AA AA AA AA |
A ili AB AB AO AB AO |
AB AB AB AB AB |
A ili AB AA AB AA AB | |
| B | IBi (BO) | O ili B BO BO OO OO |
O, A, B ili AB AB BO AO OO |
A ili AB AB AB AO AO |
O ili B BB BO BO OO |
B BB BB BO BO |
A, B ili AB AB BB AO BO |
| IBIB (BB) | B BO BO BO BO |
B ili AB AB BO AB BO |
AB AB AB AB AB |
B BB BO BB BO |
B BB BB BB BB |
B ili AB AB BB AB BB | |
| AB | IAIB (AB) | A ili B AO AO BO BO |
A, B ili AB AA AO AB BO |
A ili AB AA AA AB AB |
A, B ili AB AB AO BB BO |
B ili AB AB AB BB BB |
A, B, ili AB AA AB AB BB |
| A | B | O |
|---|---|---|
| A1 A2 A3 A4 A5 Ax Ae Aint Au Ax |
B101 (B1) B3 Bw Bx |
O01 (O1) O02 (O1v) O03 (O2) |
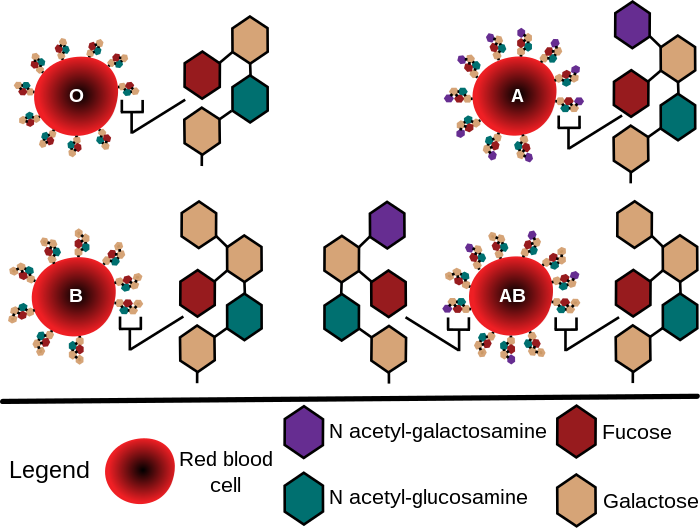
Od desetak "public" sistema krvnih grupa, u transfuziji je ABO sistem najbitnijiji.
| Fenotip | Genotip |
|---|---|
| A | IAIA ili IAI0 |
| B | IBIB ili IBI0 |
| AB | IAIB |
| O | I0I0 |
Kompatibilnost eritrocita i plazme
[uredi | uredi izvor]
Pored doniranja istoj grupi krvi, donatori O krvne grupe mogu dati krv i A, B i AB; davatelji krvi fenotipova A i B mogu dati krv osobama AB.

Pored doniranja istoj krvnoj grupi, plazma tipa AB može se dati u A, B i O; plazme A, B i AB može se dati O.
| Primatelj | Donator[1] | |||
|---|---|---|---|---|
| O | A | B | AB | |
| O | ||||
| A | ||||
| B | ||||
| AB | ||||
Isključivanje roditeljstva
[uredi | uredi izvor]| Isključenje roditeljstva po ABO sistemu krvnih grupa | ||||||
| Posmatrani par | Moguće | Isključeno | ||||
| O x O | O | A, B, AB | ||||
| O x A | O, A | B, AB | ||||
| O x B | O, B | A, AB | ||||
| O x AB | A, B | O, AB | ||||
| A x A | O, A | B, AB | ||||
| A x B | O, A, B, AB | – | ||||
| A x AB | A, B, AB | O | ||||
| B x B | O, B | A, AB | ||||
| B x AB | A, B, AB | O | ||||
| AB x AB | A, B, AB | O | ||||
Distribucija krvnih grupa ABO sistema
[uredi | uredi izvor]| Populacija | O (%) | A (%) | B (%) | AB (%) |
|---|---|---|---|---|
| Aboridžini | 61 | 39 | 0 | 0 |
| Afroamerikanci | 49 | 27 | 20 | 4 |
| Ainu (Japan) | 17 | 32 | 32 | 18 |
| Albanci | 38 | 43 | 13 | 6 |
| Američki bijelci | 45 | 40 | 11 | 4 |
| Američki starosjedioci | 79 | 16 | 4 | 1 |
| Andamanci | 9 | 60 | 23 | 9 |
| Arapi | 34 | 31 | 29 | 6 |
| Armenci | 31 | 50 | 13 | 6 |
| Austrijanci | 36 | 44 | 13 | 6 |
| Azijatski Amerikanci | 40 | 28 | 27 | 5 |
| Bantu | 46 | 30 | 19 | 5 |
| Baski | 51 | 44 | 4 | 1 |
| Belgijanci | 47 | 42 | 8 | 3 |
| Bororo (Brazil) | 100 | 0 | 0 | 0 |
| Bosanci[10] | 36 | 42 | 15 | 7 |
| Brazilci | 47 | 41 | 9 | 3 |
| Bugari | 32 | 44 | 15 | 8 |
| Burmanci | 36 | 24 | 33 | 7 |
| Burijati (Sibir) | 33 | 21 | 38 | 8 |
| Bušmani | 56 | 34 | 9 | 2 |
| Česi | 30 | 44 | 18 | 9 |
| Danci | 41 | 44 | 11 | 4 |
| Egipćani | 33 | 36 | 24 | 8 |
| Englezi | 47 | 42 | 9 | 3 |
| Eskimi (Aljaska) | 38 | 44 | 13 | 5 |
| Eskimi (Grenland) | 54 | 36 | 23 | 8 |
| Estonci | 34 | 36 | 23 | 8 |
| Etiopljani | 43 | 27 | 25 | 5 |
| Fidži | 44 | 34 | 17 | 6 |
| Filipinci | 45 | 22 | 27 | 6 |
| Finci | 34 | 41 | 18 | 7 |
| Francuzi | 43 | 47 | 7 | 3 |
| Gruzijci | 46 | 37 | 12 | 4 |
| Grci | 40 | 42 | 14 | 5 |
| Havajci | 37 | 61 | 2 | 1 |
| Hindusi (Mumbai) | 32 | 29 | 28 | 11 |
| Holanđani | 45 | 43 | 9 | 3 |
| Islanđani | 56 | 32 | 10 | 3 |
| Indijanci (pleme Crna noga - Blackfoot) | 17 | 82 | 0 | 1 |
| Indijci | 37 | 22 | 33 | 7 |
| Iranci | 38 | 33 | 22 | 7 |
| Irci | 52 | 35 | 10 | 3 |
| Holanđani | 45 | 43 | 9 | 3 |
| Italijani (Milano) | 46 | 41 | 11 | 3 |
| Japanci | 30 | 38 | 22 | 10 |
| Jevreji (Njemačka) | 42 | 41 | 12 | 5 |
| Jevreji (Poljska) | 33 | 41 | 18 | 8 |
| Južnoafrikanci | 45 | 40 | 11 | 4 |
| Kalmuki | 26 | 23 | 41 | 11 |
| Kikuju (Kenija) | 60 | 19 | 20 | 1 |
| Kinezi (Kanton) | 46 | 23 | 25 | 6 |
| Kinezi (Peking) | 29 | 27 | 32 | 13 |
| Korejanci | 28 | 32 | 31 | 10 |
| Laponci | 29 | 63 | 4 | 4 |
| Latvijci | 32 | 37 | 24 | 7 |
| Litvanci | 40 | 34 | 20 | 6 |
| Mađari | 36 | 43 | 16 | 5 |
| Malezijci | 62 | 18 | 20 | 0 |
| Maje | 98 | 1 | 1 | 1 |
| Maori | 46 | 54 | 1 | 0 |
| Navaho (Indijanci) | 73 | 27 | 0 | 0 |
| Nijemci | 41 | 43 | 11 | 5 |
| Nikobarci | 74 | 9 | 15 | 1 |
| Norvežani | 39 | 50 | 8 | 4 |
| Papuanci (Nova Gvineja) | 41 | 27 | 23 | 9 |
| Peru (Indijanci) | 100 | 0 | 0 | 0 |
| Poljaci | 33 | 39 | 20 | 9 |
| Portugalci | 35 | 53 | 8 | 4 |
| Romi (Mađarska) | 29 | 27 | 35 | 10 |
| Rumuni | 34 | 41 | 19 | 6 |
| Rusi | 33 | 36 | 23 | 8 |
| Sardinija | 50 | 26 | 19 | 5 |
| Slovaci | 42 | 37 | 16 | 5 |
| Srbija | 38 | 42 | 16 | 5 |
| Sudanci | 62 | 16 | 21 | 0 |
| Škoti | 51 | 34 | 12 | 3 |
| Šompen (Nikobar) | 100 | 0 | 0 | |
| Španci | 38 | 47 | 10 | 5 |
| Šveđani | 38 | 47 | 10 | 5 |
| Švicarci | 40 | 50 | 7 | 3 |
| Tatari | 28 | 30 | 29 | 13 |
| Turci | 43 | 34 | 18 | 6 |
| Ukrajinci | 37 | 40 | 18 | 6 |
| Vijetnamci | 42 | 22 | 30 | 5 |
| Prosjek | 43.91 | 34.80 | 16.55 | 5.14 |
| Standardna devijacija | 16.87 | 13.80 | 9.97 | 3.41 |
- A je uobičajena u Centralnoj i Istočnoj Evropi. U zemljama kao što su Austrija, Danska, Norveška i Švicarska ove grupe u ukupnoj populaciji ima oko 45–50%, dok je u Poljskoj i Ukrajini ima oko 40%. Najviše učestalosti su evidentirane u malim nesrodnim populacijama. Primjerice, oko 80% "Blackfoot" Indijanaca iz Montane imaju krvnu grupu A. Frekvencija krvne grupe A je visoka (25–55%) u cijeloj Evropi, posebno u Centralnoj i Skandinaviji. Visoka učestalost grupe A je također evidentirana i kod Aboridžina južne Australije (do 45%) i u nekim plemenima američkih Indijanaca (35%). U Evroaziji, gradijent učestastalosti krvne grupe A (i alela IA) opada prema istoku.
- Maksimalna učestalost IB je u Centralnoj Aziji i sjevernoj Indiji. Ovaj alelogen je vjerovatno odsutan kod sjevernoameričkih Indijanaca i Australskih aboridžina prije invazije bijelaca. Smatra se da ovog alela prije toga kod njih nije uopće bilo. U Evroaziji, gradijent učestastalosti krvne grupe A i (alelogena IB) raste prema istoku.
Medicinski značaj krvnih grupa ABO sistema
[uredi | uredi izvor]Medicinski značaj krvnih grupa ovog sistema antigena posebno se ogleda u transfuziji krvi, zbog moguće inkompatibilnosti antigena i antitijela donora i primatelja, kao što je prikazano u prethodnom poglavlju. Pored toga, zapažena je značajna asocijcija pojedinih krvnih grupa i prijemčivosti osoba za određene bolesti.
Najčešće je proučavana povezanost ABO fenotipova i gastričnih i duodenalnih ulcera. Pri tome je primijećena veća incidenca gastričnih i duodenalnih ulcera (30 %) kod individua O krvne grupe, u odnosu na pripadnike grupa A i B. Ta otkrića su, između ostalog, inicirala niz istraživanja mogućih veza ABO sistema krvnih grupa i nekih bolesti. U ovakvim istraživanjima primjenjivane su različite statističke metode. Kasnije su te studije proširene na relacije i nekih drugih gena i bolesti, ali od svih njih jedino je bila značajna veza između HLA i nekih bolesti. Proučavane su distribucije različitih infekcijskih, parazitskih, neoplastičnih, degenerativnih i raznih drugih oboljenja po fenotipovima ABO sistema krvnih grupa. Tako je, npr., uočena visoka korelacija između sifilisa (osobito cerebrospinalnog) i antigena A, dok su osobe krvne grupe O značajno rjeđe aficirane. Moguća povećana otpornost individua krvne grupe O na ovu bolest može se objasniti skoro potpunim odsustvom individua krvnih grupa A i B u nekim populacijama nativnih Amerikanaca, (neoindijanida), izuzimajući Eskime i neke druge sjevernoameričke skupine. Dugo se vjerovalo da je u ostali dio svijeta sifilis “uvezen“ iz Evrope nakon Kolumbovog putovanja (1492. godine). Ako je tačno da su individue krvne grupe O rezistentnije na sifilis od pripadnika ostalih krvnih grupa i da je sifilis već dugo endemična bolest u Americi, može se pretpostaviti da je prirodna selekcija eliminirala većinu individua koje nemaju ovu krvnu grupu. Nedavno su u Podsaharskoj Africi otkriveni dokazi pretpostavke da je sifilis “prenesen” i u ta područja.[11]
Neki pokazatelji nagovještavaju da i neke druge bolesti kao što su kuga, kolera i dijareja (uzrokovana patogenim E. coli) favorizuju osobe sa krvnom grupom O. Također se pretpostavlja da su i male boginje povezane sa ABO sistemom krvnih grupa, pri čemu su individue A krvne grupe rijemčivije. Neki pokazatelji indiciraju veću podložnost virozama individua krvne grupe A, uspoređujući ih sa pripadnicima B i O. Ipak, neophodno je konstatirati da odgovarajuće statističke analize dokazuju vrlo veliku heterogenost rezultata različitih istraživača, što znatno otežava jasnije zaključivanje o povezanosti ABO sistema krvnih grupa i nekih oboljenja.
Izgleda da mnoge neoplazme imaju različitu frekvenciju među ABO fenotipovima. Zabilježena je manja heterogenost rezultata nego u adekvatnim studijama infektivnih oboljenja. Neki efekti su dosta specifični, kao npr., rjeđa opažanja maligne neoplazme pljuvačne žlijezde, jednjaka i pilorusa kod osoba krvne grupe O, koje su, izgleda, rezistentnije nego pripadnici grupa A i B. Pri tome, ostatak gastrointestinalnog trakta nije diferencijalno inficiran s obzirom na pripadnost ABO fenotipovima. Postoje i dokazi da su osobe krvne grupe O rezistentnije na adenokarcinome.[11][12]
Među ostalim oboljenjima, dijabetes ispoljava nešto veću prijemčivost za A i B u odnosu na O krvnu grupu, perniciozna anemija je 4–5, a miopija dva i po puta češća kod A i B u usporedbi sa O. Zabilježena je i veća stopa reumatičnih i srčanih oboljenja, tromboze, embolije i ciroze jetre kod osoba sa A i B krvnom grupom u odnosu na O, a suprotni nalazi su registrirani za reumatoidni artritis.
Relacije glavnih krvnih grupa ABO sistema i nekih bolesti
| ||||||
| Pogođena | Referentna | Bolest | ||||
| A | O | Kanceri | ||||
| A ("ne–O") |
O | |||||
| A i B | O | |||||
| O | "ne-O" | Ulkusi | ||||
Rh sistem krvnih grupa
[uredi | uredi izvor]Antigeni i genetika Rh sistema
[uredi | uredi izvor]Prisustvo i karakterizacija osnovnih krvnih grupa Rh (rezus) sistema definirani su u vezi sa njihovom ulogom u pojavi hemolitske bolesti fetusa i novorođenčeta (erythroblastosis fetalis, morbus haemolyticus neonatorum), koja se manifestuje u intrauterinoj ili perinatalnoj destrukciji (aglutinaciji) eritrocita ploda antitijelima za odgovarajuću antigenu supstancu . U takvim slučajevima crvena krvna zrnca djeteta sadrže rezus protein, koji se redovno susreće kod rezus majmuna, ali i kod oko 85% pripadnika kavkazoidnog stanovništva. Prva reakcija antigen – antitijelo u ovom sistemu varijacija detektirana je nakon eksperimentalnog injiciranja krvi rezus makaki majmuna (Macaca rhesa = Macacus rhesus = Macacus mulattus) u cirkulacijski sistem kunića. Prisustvo rezus proteina na eritrocitima označava se kao Rh+’’, a njegovo odsustvo kao Rh–.
Antigeni Rh sistema krvnih grupa predstavljaju proizvod dva čvrsto vezana i visoko homologna gena: RHD i RHCE (zajednički se nazivaju RH30 ili RHCED). RhD nosi antigen D koji je najsnažniji imunogen krvnih grupa ali je također i odsutan u relativno velikom dijelu populacije (tj. Rh– fenotip) što je rezultat delecije RHD gena ili drugog tipa promjena. RHCE postoji u četiri alelne forme i svaki alel određuje ekspresiju dva antigena u kombinacijama Ce, ce, cE ili CE (RHCE je zajednički naziv za četiri alela). Geni RHD i RHCE se sadrže 10 egzona i protežu se duž 75 kb DNK sekvence. Za antigenu aktivnost Rh neophodno je formiranje kompleksa sa Rh50 glikoproteinom, produktom još jednog funkcionalnog gena RHAG (naziva se još i RH50. RHAG je slično organiziran u 10 egzona i sličan je RHCED u 36% sekvence ali se nalazi na zasebnom lokusu. Dok su alteracije RHAG lokusa relativno rijetke, lokus RHCED raspolaže širokom paletom alelnog diverziteta na nivou populacija.
Open reading frames (ORF), tj. funcionalni okviri za transkripciju, RHCE i RHD su suprotno orijentirani – njihovi 3' krajevi su okrenuti jedan prema drugome i razdvojeni dijelom DNK (veličine 30 kb) u kojem je smješten gen SMP1. Osim toga, na 5’ i 3’ krajevima RHD nalaze se i dvije visoko homologe sekvence (veličine 9 kb), koje se nazivaju Rhesus kutije (Rhesus boxes). Pretpostavlja se da do delecije RHD, koja se često susreće u populaciji, dolazi usljed inekvalne homologe rekombinacije ograničene na ove sekvence.
Produkti RHCED i RHAG su integralni membranski proteini sa sličnom 12–transmembranskom helikoidnom topologijom. Rh 30 polipeptidi su palmitolirani i njihove epitope definira pet specifičnih aminokiselina u ekstracelularnim petljama. Rh50 je N–glikoziliran u jednom mjestu – Asn37. Dokazano je postojanje dva ne-eritrocitna homologa RHAG nazvana RHBG i RHCG. Imaju visok nivo homologije, ali njihovi načini ekspresije nisu identični.
Genski lokusi Rh sistema antigena nalaze na kratkom kraku hromosoma 1: RHD i RHCE na 1p36.2–p34, a lokus za RHAG (za Rezus asocirani glikoprotein) je na kratkom kraku hromosoma 6: 6p21.1–p11.[13]
Ako u eritrocitima majke nedostaje takav protein (tj. ako je rezus–negativan: Rh–) imuni odgovor njenog organizma na rezus–pozitivne Rh+ krvne ćelije ploda je produkcija odgovarajućih antitijela. Senzibilizacija sa identičnim posljedicama moguća je i nakon transfuzije krvi rezus–pozitivnog donatora rezus–negativnom recipijentu. Studioznija istraživanja ovog značajnog sistema kvalitativne varijacije bila su posebno stimulirana činjenicom da je neposredno nakon otkrića dimorfizma prisustvo – odsustvo rezus proteina u ljudskim eritrocitima (Landsteiner, Wiener 1940) registrovano da se u preko 90% slučajeva hemolitičke bolesti radi o inkopatibilnosti između rezus–negativne majke i rezus–pozitivnog ploda (Levine 1941). Također je konstatirano da se imunološke komplikacije i hemolitska bolest obično javljaju u takvim konstelacijama druge i narednih trudnoća (odnosno nakon višekratnih transfuzija krvi) jer producirana količina antitijela u prvoj trudnoći najčešće nije dostatna za ozbiljnije posljedice. Prema tome, za razliku od antitijela ABO sustava, koja su redovno prisutna u normalnom serumu, produkcija Rh antitijela se javlja kao imuni odgovor na unošenje odgovarajućih antigena u organizam. Inače, osim krvi (eritrocita), Rh antigeni se nalaze i u ostalim tkivima i nekim izlučevinama.
Prema najjednostavnijoj (i prvoformuliranoj) hipotezi o prirodi nasljeđivanja krvnih grupa rezus sistema, baziranoj na proučavanju distribucije dva osnovna fenotipa u potomstvu pojedinih tipova braka, oni su izraz interakcije samo dvije genske varijante (R, r) jednog autosomalnog lokusa sa kompletnom funkcionalnom dominacijom alela koji determinira prisustvo odgovarajuće (R) proteinske supstance u eritrocitima. Iako su kasnija istraživanja pokazala da se ovdje radi o daleko složenijem sistemu kvalitativne varijacije (i njegove genetičke kontrole), u široj upotrebi se još uvijek održava njegova kategorizacija na dvije pomenute glavne grupe (kao pojednostavljeni izraz savremenih hipoteza o njihovoj feno–genotipskoj prirodi). Tako su dosadašnja proučavanja pokazala da klasični dominantni fenotip (R) obuhvata najmanje sedam osnovnih antigenih supstanci, tj. da se ovo svojstvo javlja u ne manje od osam glavnih grupno–specifičnih varijanti, od kojih se neke diferenciraju i u više subkategorija . Teorijsko objašnjenje konstatiranih nalaza suvremena genetika nalazi u dvije (gotovo jednako) prihvatljive hipoteze. Prema multiplo–alelnoj teoriji (Wiener; popularnija u referentnoj američkoj literaturi) ovaj sistem antigene varijacije određuje serija od osam multiplih alela odgovarajućeg lokusa, koji se ispoljavaju u isto toliko antigenih specifičnosti (r, R1, R2, R0, R', R , RyRz, od kojih R1), R2 i r svojom ukupnom frekvencijom čine ogromnu većinu (preko 95%), dok su ostali "rijetki" ili "veoma rijetki".
U identifikaciji fenotipova i genotipova Rh sistema:
- svako od poznatih antitijela aglutinira više nego jedan antigeni tip,
- svaki antigen reaguje sa više nego jednim specifičnim antitijelom, i
- svaki set antigenih osobenosti (R1, r, itd.) nasljeđuje se kao jedinstvena cjelina.
Jedna od neposrednih posljedica tog fenomena je i konstatirana učestalost ovih antigena (koja nije u suglasnosti sa hipotezom alternativne varijacije ovog svojstva). Imajući to u vidu, teorija gen–kompleksa (Fisher; šire prihvaćena u Evropi) minimum prihvatljivih objašnjenja genetičkih osnova ovog sustava krvnih grupa nalazi u pretpostavci da individualni Rh fenotip kontroliraju tri alelna para iz odgovarajuće serije blisko vezanih autosomnih lokusa (hromosoma 1 humane garniture) sa različitim haploidnim kombinacijama – haplotipovima. Općenito gledano, a prema standardnoj definiciji, haplotip je set svih alela različitih lokusa jednog hromosoma, koji su odgovorni za ekspresiju posmatranog svojstva. Pošto je među njima mogućnost rekombinacija i teorijski zanemarljiva, u Rh sistemu krvnih grupa svaki tročlani set alela se praktično nasljeđuje kao cjelovita funkcionalna jedinica (kao u primjerima jednostavnog monogenskog nasljeđivanja). Kako u parovima alela C – c, D – d i E – e nema odnosa dominantnosti, prisustvo svakog od njih u genotipu redovno se i fenotipski manifestuje kao prisustvo odgovarajućeg antigena u eritrocitima. Prema tome u klasičnom rezus negativnom fenotipu koji ne sadrži ni C, ni D, ni E antigen također su prisutne grupno–specifične supstance (značajno slabije antigene aktivnosti) ovog sustava, tj. u ovoj nomenklaturi odgovara mu kombinacija cde. Kao veoma rijetki aleli (i odgovarajući antigeni “specifikumi”) registrirani su Cw, Cx, Cu, Cv, Du, Dw, Eu, Ev, i dr. Antigenost opada u smjeru D–C–c–E–Cw–e.
Međusobnim kombinovanjem 8 haplotipova tri pomenuta lokusa nastaje 36 različitih individualnih genotipova (teorijski izvodivih iz hibridizacije genotipa CDE/cde); respektujući, međutim, njihov genski sastav, taj broj se svodi na 28, budući da su neki od njih (CDE/cde i Cde/cDE, naprimjer) u tom pogledu ekvivalentni. Suglasno tome, a prema ovom viđenju prirode nasljeđivanja krvnih grupa rezus sistema, broj mogućih kombinacija roditeljskih genotipova (tipova braka) iznosi 666. Alelni tripleti Cde (R1), cde (r) i Cde (R2) čine ukupno čine preko 95% genskog fonda evropskog stanovništva.
| Simboli i relativna učestalost genetičkih determinatora Rh krvnih grupa u evropskom stanovništvu (prema dvije rivalske teorije) | ||||||
| Gen-kompleks (Wiener) | Multipli alelizam (Fisher – Race) | Učestalost (%) | ||||
| r | cde | 39,0 | ||||
| R1 | CDe | 42,0 | ||||
| R2 | cDE | 14,0 | ||||
| R0 | cDe | 2,5 | ||||
| R, | Cde | 1,0 | ||||
| R,, | cdE | 1,0 | ||||
| Ry | CdE | vrlo rijetko | ||||
| CDE | Rz | >0,25 | ||||
| Relativna frekvencija (%) antigena Rh sistema u evropeidnom stanovništvu | ||||||
| Gen-kompleks (Wiener) | Multipli alelizam (Fisher – Race) | Učestalost (%) | ||||
| Rh | C | 70 | ||||
| Rh0 | D | 85 | ||||
| Rh | E | 30 | ||||
| hr, | c | 80 | ||||
| Hr0 | d | 60 | ||||
| Hr,, | e | 96 | ||||
| Teorijska distribucija Rh krvnih grupa u potomstvu mogućih (genotipskih) tipova braka (%) | ||||||
| Roditelji | Potomstvo | |||||
| RhRh x RhRh | RhRh : 100 | |||||
| RhRh x Rhrh | RhRh 50 + Rhrh: 50 | |||||
| Rhrh x Rhrh | RhRh: 25 + Rhrh: 50 + rhrh: 25 | |||||
| RhRh x rhrh | Rhrh: 100 | |||||
| Rhrh x rhrh | Rhrh: 50 + rhrh: 50 | |||||
| rhrh x rhrh | rhrh: 100 | |||||
| Relativne frekvencija Rh– krvne grupe u odabranim populacijama (%) | ||||||
| Populacija | Rh–(%) | |||||
| Australija | 19 | |||||
| Austrija | 18 | |||||
| Belgija | 15,3 | |||||
| Bosna i Hercegovina | 15 | |||||
| Danska | 16 | |||||
| Finska | 13 | |||||
| Francuska | 15 | |||||
| Holandija | 15,3 | |||||
| Hong Kong | <1,5 | |||||
| Irska | 16 | |||||
| Južna Koreja | 0,35 | |||||
| Kanada | 4,9 | |||||
| Novi Zeland | 18 | |||||
| Njemačka | 15 | |||||
| Poljska | 15 | |||||
| SAD | 15 | |||||
| Švedska | 16 | |||||
| UK | 17 | |||||
| Evropski prosjek | 15 | |||||
Medicinski značaj Rh sistem krvnih grupa
[uredi | uredi izvor]
Rh+ - crveno
Rh- - plavo

Kada je riječ o Rh sistemu eritrocitnih antigena, oni su, ustvari, i detektirani u pokušajima objašnjenja uzroka prenatalne i perinatalne hemolitičke anemije (erythroblastosis fetalis) u slučajevima kada Rh- majka nosi plod krvne grupe Rh+. To se dešava zbog inkompatibilnosti seruma majke i Rh antigena djeteta. Iz istog razloga, osobe krvne grupe Rh-, prilikom transfuzije, ne smiju primati krv Rh+. Također je zabilježeno da su osobe sa rezus–antigenom D i MNS antigenom N naročito podložniji malim boginjama.
Međusobna kompatibilnost kombinacija antigena ABO i Rh sistema[14][15]
[uredi | uredi izvor]| Primatelj[1] | Donator[1] | |||||||
|---|---|---|---|---|---|---|---|---|
| O- | O+ | A- | A+ | B- | B+ | AB- | AB+ | |
| O- | ||||||||
| O+ | ||||||||
| A- | ||||||||
| A+ | ||||||||
| B- | ||||||||
| B+ | ||||||||
| AB- | ||||||||
| AB+ | ||||||||
Distribucija kombinacija glavnih krvnih grupa ABO i Rh sistema
[uredi | uredi izvor]| Država | O+ | A+ | B+ | AB+ | O− | A− | B− | AB− |
|---|---|---|---|---|---|---|---|---|
| Austrija[16] | 30 | 33 | 12 | 6 | 7 | 8 | 3 | 1 |
| Australija[17] | 40 | 31 | 8 | 2 | 9 | 7 | 2 | 1 |
| Belgija[18] | 38.1 | 34 | 8.5 | 4.1 | 7 | 6 | 1.5 | 0.8 |
| Bosna i Hercegovina[11] | 31 | 36 | 12 | 6 | 5 | 7 | 2 | 1 |
| Danska[19] | 35 | 37 | 8 | 4 | 6 | 7 | 2 | 1 |
| Finska[20] | 27 | 38 | 15 | 7 | 4 | 6 | 2 | 1 |
| Francuska[21] | 36 | 37 | 9 | 3 | 6 | 7 | 1 | 1 |
| Holandija[22] | 39.5 | 35 | 6.7 | 2.5 | 7.5 | 7 | 1.3 | 0.5 |
| Hong Kong, Kina[23] | 40 | 26 | 27 | 7 | <0.3 | <0.3 | <0.3 | <0.3 |
| Irska[24] | 47 | 26 | 9 | 2 | 8 | 5 | 2 | 1 |
| Južna Koreja[25] | 27.2 | 35.1 | 26.1 | 11.3 | 0.1 | 0.1 | 0.1 | 0.05 |
| Kanada[26] | 39 | 36 | 7.6 | 2.5 | 7 | 6 | 1.4 | 0.5 |
| Novi Zeland[27] | 38 | 32 | 9 | 3 | 9 | 6 | 2 | 1 |
| Njemačka[28] | 35 | 37 | 9 | 4 | 6 | 6 | 2 | 1 |
| Poljska[29] | 31 | 32 | 15 | 7 | 6 | 6 | 2 | 1 |
| SAD[30] | 37.4 | 35.7 | 8.5 | 3.4 | 6.6 | 6.3 | 1.5 | 0.6 |
| Švedska[31] | 32 | 37 | 10 | 5 | 6 | 7 | 2 | 1 |
| Ujedinjeno Kraljevstvo[32] | 37 | 35 | 8 | 3 | 7 | 7 | 2 | 1 |
MN(Ss) sistem krvnih grupa
[uredi | uredi izvor]Antigeni i genetika MN(Ss) sistema
[uredi | uredi izvor]MN(Ss)sistem krvnih grupa obuhvata takva svojstva humanih eritrocita, koja se, između ostalog, manifestuju se i u tome što alternativno posjeduju samo jednu ili istovremeno obje antigene supstance označene sa M i N. Na osnovu toga, ljudsku populaciju je moguće podijeliti na pripadnike M, N i MN krvne grupe ovog sistema.[33][34]
Prisustvo ili odsustvo antigena M i N kod čovjeka je detektirano nakon injiciranja prepariranih eritrocita različitih osoba u krvotok kunića, čija se imunog odgovora ispoljava u produkciji specifičnih antitijela za homologe antigene tvari M i N. Paralelni kontakt testiranih crvenih krvnih zrnaca (bilo koje osobe) sa ova dva specifična antitijela može imati tri alternativne varijante ishoda; aglutinacija sa anti–M, sa anti–N i sa oba, na osnovu čega se i identificiraju krvne grupe M, N i MN. Prema tome, grupno–specifična antitijela (anti–M i anti–N) ovog sistema u organizmu pokusnih kunića nastaju kao imuni odgovor na odgovarajuću (M i N) proteinsku supstancu, a opisana aglutinacijska reakcija dokazuje da se u MN eritrocitima nalaze obje antigene specifičnosti ovog sistema. MN antigeni redovno su prisutni u humanim crvenim krvnim ćelijama, dok odgovarajuća antitijela u normalnoj krvi nisu registrirana. Međutim, u slučajevima heterospecifičnih transfuzija N–davaoca i M–primaoca krvi i obrnuto, stvaraju se specifična antitijela slabe reaktivne sposobnosti, koja ne perzistiraju tako dugo da bi izazvala ozbiljnije komplikacije u eventualnim narednim transfuzijskim zahvatima. Ako se tome doda činjenica da osobe MN fenotipa uopće ne produciraju anti–M i anti–N tjelašca (budući da proteini M i N za njih nisu strani), postaje jasno zašto je trasfuzijski značaj krvnih grupa MN sistema praktično zanemarljiv. Međutim, aloimunizacija aloantigenima MNSs sistema ipak može ponekad izazvati hemolitsku bolest novorođenčeta.
| Aglutinogeni i aglutinini MN sistema krvnih grupa | ||||||
| Eritrociti (Antigeni) | Test serum anti–M | Test serum anti–N | ||||
| M | + | – | ||||
| N | – | + | ||||
| MN | + | + | ||||
- + Aglutinacija
- – Nema aglutinacije
Antigeni MNSs sistema su glikoforinske (GP) prirode – membranski proteini tipa I. Eritrocitarne membrane sadrže dvije vrste ovih supstanci: glikoforin A (GP–A, molekulske mase 31 kDa) i glikoforin B (GP–B, molekulske mase 11 kDa), a diferencijacija specifičnosti im počiva na osobenostima aminosekvence na pozicijama 1 i 5 (varijanta A), odnosno 29 (varijanta B). Glikoforin A daje aloantigenu svojstvenost M u kojoj se na poziciji 1 javlja serin, a na poziciji 5 – glicin. Antigen N na poziciji 1 ima leucin, a na poziciji 5 je glutaminska kiselina. Glikoforin B daje aloantigene S (metionin na poziciji 29) i s (treonin na poziciji 29).[35]
Uloga GPA i GPB još uvijek nije razjašnjena. Izgleda da do ekspresije GPE, još jednog člana ove porodice gena, ne dolazi u normalnim fiziološkim uvjetima.Također je poznato da odsustvo GPA i/ili GPB iz eritrocita, što je dokumentovano u nekim varijantama eritrocita u homozigotnom stanju (Mk, En(a), S–s–), ne dovodi do značajnih fizioloških abnormalnosti.
Pripadnost krvnim grupama M, N i MN isključivo je genetički određena, tj. na fenotipsko ispoljavanje njihovih genetičkih determinatora spoljni činioci nemaju nikakvog uticaja. Na odgovarajućem (autosomalnom) genskom lokusu (alternativno) se javljaju dva alela (LM i LN), među kojima nema odosa dominantnosti – recesivnosti pa se prisustvo svakog od njih u individualnoj genetičkoj konstituciji fenotipski ispoljava u sintezi odgovarajuće antigene supstance, neovisno o aktivnosti druge alelne varijante (ili homospecifičnog parnjaka na drugom homologom hromosomu). Prema tome, tri moguće genotipske kombinacije ovih alela (LMLM, LMLN i LNLN) daju isto toliko fenotipskih ekspresija (M, N i MN). Mogući tipovi braka i očekivana distribucija genotipova (i fenotipova) u njihovom potomstvu (prema važećoj teoriji o prirodi nasljeđivanja krvnih grupa MN sistema), prikazani su u tabeli 3.15.
Osim M i N supstanci, pod različitim nazivima je opisan niz antigenih varijanti koje ispoljavaju njima srodna imunološka i druga specifična svojstva. Tako je u ljudskom serumu najprije registrirano jedno specifično antitijelo za koje se kasnije i dokazalo da odgovara eritrocitarnom antigenu S te da je njegovo prisustvo genetički kontrolirano sa genskog lokusa koji je blisko vezan za odgovarajuću MN poziciju. Imajući u vidu zanemarljivu učestalost rekombinacija između ova dva lokusa, njihove alelne kombinacije (MS, Ms, NS i Ns) se nasljeđuju i tretiraju kao integralne nasljedne jedinice (haplotipovi dvojakog [fenotip]skog izraza), koje se u sukcesivnim generacijama posmatranih rodoslova ponašaju kao aleli jednostavnih monogenskih svojstava.
MN i Ss aleli se nalaze na blisko vezanim lokusima hromosoma 4; preciznije, klaster MN(S) gena, približne veličine 350 kb (5,–GYPA-GYPB–GYPE-3,), lociran je na njegovom dugom kraku: 4q28-q31.
Genetička kontrola ovog sistema antigena odvija se, dakle, putem dva alelogena sa hromosoma 4 ljudske hromosomske garniture, koji kodiraju (glikoforinA i glikoforin B). U okviru ovog sistema opisano je čak 46 antigena.[36][37][38][39], među kojima su najznačajniji M, N, S, s i U. Respektirajući tu činjenicu, u MNS sistemu se javlja šest mogućih fenotipova: MS, Ms, NS, Ns, MNS i MNs.
Distribucija krvnih grupa MN(S) sistema
[uredi | uredi izvor]Najćešći su M+ i N+ eritrociti (75% ), a M + N + je najčešći genotip (50% ). S antigen je relativno čest (~ 55% ), a e još frekventniji (~ 89% ). U antigen ima također visoku učestalost, a javlja se u više od 99,9% osoba. U je prvobitno bila skraćenica od "Universal", iako to nije slučaj. Kod RBCs negativnih eritrocita može se naći u starosjedilačkom stanovnišvu Afrike .
Kada je riječ o MN(S) sistemu krvnih grupa, primijećena je određena razlika u distribuciji alela i haplotipova. M i N aleli su regularno distribuirani, ali je alel s rijetka varijanta u Jugoistočnoj Aziji, a gotovo ga i nema u Australiji. Alel Su je gotovo isključivo vezan za Afriku. Od četiri haplotipa, dva gotovo da i ne postoje u Australiji (MS i NS), a MS je rijedak u Novoj Gvineji i na nekim pacifičkim otocima.
| Distribucija glavnih krvnih grupa MN sistema u odabranim populacijama[40] | ||||||
| Populacija | Uzorak | M (%) | N (%) | M (%) | ||
| Aboridžini | 372 | 3 | 30 | 67 | ||
| Ainu (Japan) | 504 | 18 | 50 | 32 | ||
| Aleuti | 132 | 68 | 29 | 3 | ||
| Arapi (Beduini) | 206 | 37 | 49 | 14 | ||
| Armeni | 339 | 34 | 45 | 21 | ||
| Baski (Španija) | 91 | 23 | 52 | 25 | ||
| Belgijanci | 3.100 | 29 | 50 | 21 | ||
| Bosanci | 3.204 | 30 | 50 | 20 | ||
| Danci | 2.023 | 29 | 50 | 21 | ||
| Egipćani | 419 | 26 | 53 | 21 | ||
| Eskimi | 569 | 83 | 16 | 1 | ||
| Estonci | 310 | 35 | 50 | 15 | ||
| Filipinci | 382 | 26 | 50 | 24 | ||
| Finc | 1.050 | 37 | 47 | 16 | ||
| Francuzi | 400 | 33 | 46 | 21 | ||
| Indonežani | 296 | 30 | 46 | 24 | ||
| Irci | 399 | 30 | 47 | 23 | ||
| Italijani | 300 | 32 | 48 | 20 | ||
| Japanci | 1.100 | 32 | 47 | 21 | ||
| Kinezi | 1.029 | 33 | 49 | 18 | ||
| Mađari | 624 | 33 | 48 | 19 | ||
| Navaho (Indijanci) | 361 | 85 | 14 | 1 | ||
| Sirijci | 306 | 31 | 52 | 17 | ||
| Škoti | 456 | 35 | 48 | 17 | ||
| Šveđani | 1.200 | 36 | 47 | 17 | ||
| Ukrajinci | 310 | 36 | 44 | 20 | ||
| Ukrajinci | 310 | 36 | 44 | 20 | ||
| Velšani | 192 | 31 | 55 | 14 | ||
Medicinski značaj krvnih grupa MN(Ss) sisetema
[uredi | uredi izvor]Suvremena istraživanja povezanosti MNS sistema krvnih grupa i mentalnih oboljenja ukazuju na mogućnost asociranosti lokusa na hromosomu 4 (4q28–q31.1) sa nasljednom prijemčivosti za bipolarni poremećaj (BD – bipolar disorder). Naime, ispostavilo se da je broj individua genotipa LNLN (N fenotip) u ispitivanoj skupini pacijenata sa bipolarnim poremećajem ličnosti signifikantno manji nego u kontroli (osobe ostalih fenotipova MN sistema krvnih grupa). Također je zabilježeno da su osobe sa rezus–antigenom D i MNS antigenom N podložniji malim boginjama.
Izvjesni problemi usljed inkopatibilnosti majka – plod rijetko su zabilježeni i prema antigenima ABO i MN sistema.
Reference
[uredi | uredi izvor]- ^ McCullough J (2011): Transfusion medicine. Published Online: 8 NOV 2011 04:17AM EST, Print ISBN 9781444337051 Online ISBN 9781444398748
- ^ Mourant A. E. et al. (1976): The human blood groups and other polymorphisms. Oxford University Press, London.
- ^ Reid M. E., Lomas-Francis C. (2004): The Blood Group Antigen Facts Book. Second ed., Elsevier Academic Press New York.
- ^ Daniels G. (2002): Human Blood Groups, Second ed. Blackwell Science.
- ^ Ridley M. (2001): Genom. Izvori, Zagreb, ISBN 953-203-091-3.
- ^ OMIM
- ^ http://www.omim.org/
- ^ http://www.ncbi.nlm.nih.gov/books/NBK2267/
- ^ https://upload.wikimedia.org/wikipedia/commons/7/78/ABO_Blood_type.jpg
- ^ Hadžiselimović R., Pojskić N. (2005): Uvod u humanu imunogenetiku. Institut za genetičko inženjerstvo i biotehnologiju (INGEB), Sarajevo.
- ^ a b c Hadžiselimović R., Pojskić N. (2005): Uvod u humanu imunogenetiku. Institut za genetičko inženjerstvo i biotehnologiju (INGEB), Sarajevo, ISBN 9958-9344-3-4.
- ^ Hadžiselimović R. (2005): Bioantropologija – Biodiverzitet recentnog čovjeka. Institut za genetičko inženjerstvo i biotehnologiju (INGEB), Sarajevo, ISBN 9958-9344-2-6.
- ^ http://www.ncbi.nlm.nih.gov/books/NBK2269/BO
- ^ a b "RBC compatibility table". American National Red Cross. 2006. Arhivirano s originala, 4. 8. 2007. Pristupljeno 24. 12. 2006. Nepoznati parametar
|month=zanemaren (pomoć) - ^ a b Blood types and compatibility Arhivirano 19. 4. 2010. na Wayback Machine bloodbook.com
- ^ "Austrian Red Cross - Blood Donor Information". Arhivirano s originala, 13. 2. 2008. Pristupljeno 26. 5. 2008.
- ^ Blood Types - What Are They?, Australian Red Cross
- ^ "Rode Kruis Wielsbeke - Blood Donor information material". Arhivirano s originala, 22. 4. 2008. Pristupljeno 26. 5. 2008.
- ^ "Frequency of major blood groups in the Danish population". Arhivirano s originala, 17. 8. 2009. Pristupljeno 26. 5. 2008.
- ^ "Suomalaisten veriryhmäjakauma". Arhivirano s originala, 26. 10. 2019. Pristupljeno 26. 5. 2008.
- ^ "Les groupes sanguins (système ABO)". Centre Hospitalier Princesse GRACE - Monaco (jezik: French). C.H.P.G. MONACO. 2005. Arhivirano s originala, 25. 12. 2018. Pristupljeno 27. 12. 2006.CS1 održavanje: nepoznati jezik (link)
- ^ Blood bank Sanquin - Blood Donor information material
- ^ "Blood Donation, Hong Kong Red Cross". Arhivirano s originala, 21. 2. 2008. Pristupljeno 26. 5. 2008.
- ^ "Arhivirana kopija". Arhivirano s originala, 3. 5. 2008. Pristupljeno 26. 5. 2008.CS1 održavanje: arhivirana kopija u naslovu (link)
- ^ "Statistics on annual blood donations 2002-2007, Gyung Nam Blood Center". Arhivirano s originala, 11. 3. 2008. Pristupljeno 26. 5. 2008.
- ^ "Types & Rh System, Canadian Blood Services". Arhivirano s originala, 4. 11. 2014. Pristupljeno 26. 5. 2008.
- ^ New Zealand Blood Service
- ^ de:Blutgruppe#Häufigkeit der Blutgruppen
- ^ Regionalne Centrum Krwiodawstwa i Krwiolecznictwa we Wrocławiu
- ^ "Blood Types in the U.S." Arhivirano s originala, 12. 6. 2010. Pristupljeno 26. 5. 2008.
- ^ "Frequency of major blood groups in the Swedish population". Arhivirano s originala, 24. 11. 2010. Pristupljeno 26. 5. 2008.
- ^ "Frequency of major blood groups in the UK". Arhivirano s originala, 11. 10. 2009. Pristupljeno 26. 5. 2008.
- ^ Mark E., Brecher E. Eds (2005): AABB Technical Manual, 15th edition. Bethesda, MD, ISBN 1-56395-196-7
- ^ Denise M. Harmening (1999): Modern Blood Banking and Transfusion Practices. F.A. Davis Company, Philadelphia, PA.
- ^ http://www.ncbi.nlm.nih.gov/books/NBK2274/
- ^ Daniels G, Flegel WA, Fletcher A, et al. International Society of Blood Transfusion Committee on Terminology for Red Cell Surface Antigens: Cape Town Report. Vox Sang 2007; 92: 250-3.
- ^ Poole J., Daniels G. (2007): Blood group antibodies and their significance in transfusion medicine. Transfus Med Rev 2007; 21: 58-71.
- ^ Daniels G. (2002): Human Blood Groups – 2nd Ed. Blackwell Science, Oxford, London.
- ^ ISBT Committee on Terminology for Red Cell Surface Antigens. "Table of blood group antigens within systems". International Society for Blood Transfusion. Arhivirano s originala, 18. 8. 2011. Pristupljeno 24. 1. 2010.
- ^ Boyd W. F. (1950): Genetics and the races of man - An introduction to modern physical anthropology. Little, Brown and Company, Boston.
Vanjski linkovi
[uredi | uredi izvor]- BGMUT "Blood Group Antigen Gene Mutation Database" u National Center for Biotechnology Information (NCBI), NIH ima detalje gena i proteina, i njihove varijacije, koje su nadležne za tipove krvi
- http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=1082436 Arhivirano 1. 8. 2013. na: Archive.today
- https://web.archive.org/web/20051102025629/http://www.bloodbook.com/type-sys.html, Blood typing systems other than ABO, BloodBook.com
- https://web.archive.org/web/20060926110321/http://www.lifeshare.org/facts/raretraits.htm, Blood Facts, Rare Traits, LifeShare Blood Centers
- http://anthro.palomar.edu/vary/vary_3.htm Arhivirano 11. 12. 2009. na Wayback Machine, Modern Human Variation: Distribution of Blood Types, O'Neil D.: Behavioral Sciences Department, Palomar College, San Marcos, California. *http://web.archive.org/web//
- http://anthro.palomar.edu/vary/vary_3.htmhttp://web.archive.org/web//[mrtav link]
- http://anthro.palomar.edu/vary/vary_3.htm Arhivirano 11. 12. 2009. na Wayback Machine.
- https://web.archive.org/web/20100304222457/http://www.bloodbook.com/world-abo.html, Racial and Ethnic Distribution of ABO Blood Types - BloodBook.com, Blood Information for Life, authorlinkbloodbook.com