Lipid
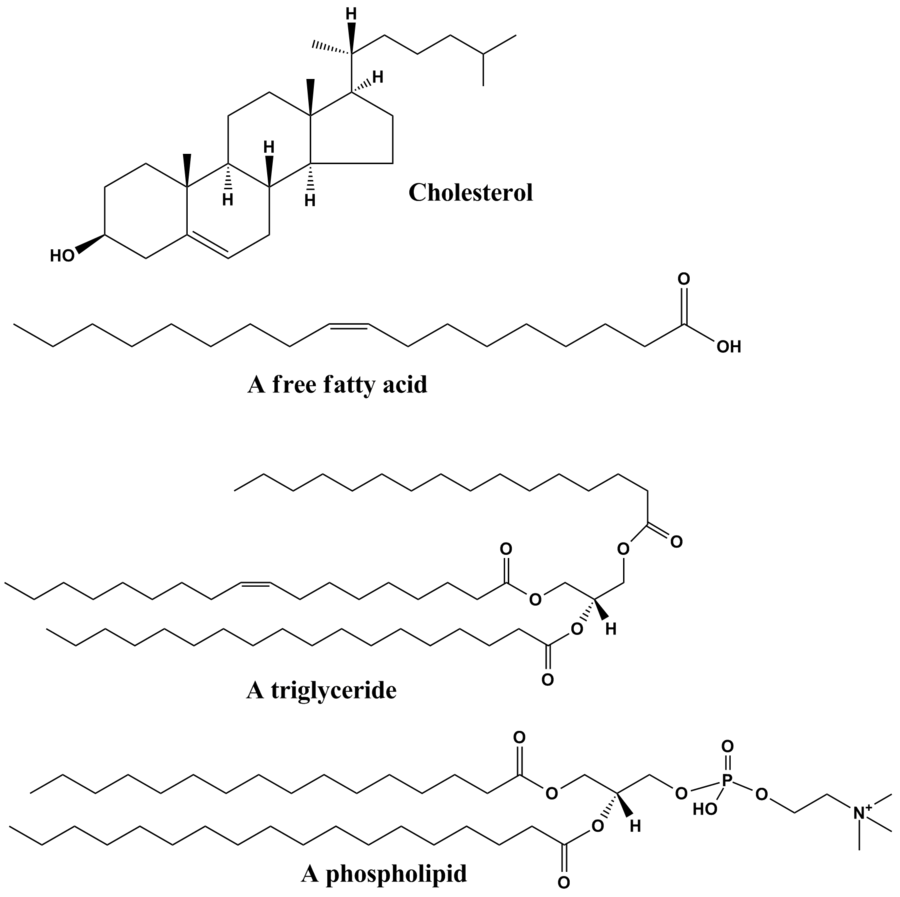
Lipider er en gruppe af naturligt forekommende molekyler, der omfatter fedtstoffer, voks, steroler, fedtopløselige vitaminer (såsom vitamin A, D, E og K), monoglycerider, diglycerider, triglycerider, fosfolipider og andre. Lipiders centrale biologiske funktioner omfatter at opbevare energi, signalere, og fungere som en strukturel komponent i cellemembraner.[4][5] Lipider anvendes indenfor kosmetik og fødevareindustrien, samt indenfor nanoteknologi.[6]
Lipider kan bredt defineres som hydrofobe eller amfifile små molekyler; nogle lipiders amfifile natur tillader dem at danne strukturer såsom vesikler, multilamellare/unilamellare liposomer, eller membraner i et vandigt miljø ved selvsamling. Biologiske lipider opstår fuldstændigt eller delvist fra to forskellige typer biokemiske underenheder eller "byggesten": ketoacyl og isopren-grupper.[4] Ved at bruge denne tilgang kan man opdele lipider i otte kategorier: fedtsyrer, glycerolipider, glycerofosfolipider, sphingolipider, sakkarolipider og polyketider (fra kondensering af ketoacyl-underenheder); samt sterollipider og prenollipider (fra konsensering af isopren-underenheder).[4]
Selvom begrebet lipid somme tider bruges som synonym for fedtstof er fedtstoffer en undergruppe af lipider kaldet triglycerider. Lipider omfatter molekyler såsom fedtsyrer og deres derivater (heriblandt tri-, di-, monoglycerider og fosfolipider), såvel som andre sterol-indeholdende metabolitter såsom kolesterol.[7] Selvom mennesker og andre pattedyr bruger diverse biosyntetiske veje til at nedbryde og syntetisere lipider, er der nogle essentielle lipider, der ikke kan skabes på denne måde og skal opnås gennem kosten.
Lipidkategorier
[redigér | rediger kildetekst]Fedtsyrer
[redigér | rediger kildetekst]Fedtsyrer, eller fedtsyreremanenser når de er en del af et lipid, er en forskelligartet molekylegruppe syntetiseret af kæde-forlængelse af en acetyl-CoA-primer med malonyl-CoA- eller metylmalonyl-CoA-grupper i en proces kaldet fedtsyresyntese.[8][9] De består af en carbonhydrid-kæde, der slutter i en carboxylsyre-gruppe; dette arrangement giver molekylet en polær, hydrofil ende, og en ikke-polær, hydrofob end, der er uopløselig i vand. Fedtsyrestrukturen er en af de mest grundlæggende kategorier af biologiske lipider, og bruges ofte som en byggesten til mere strukturelt komplekse lipider. Carbonkæden, der typisk er mellem fire og 24 carboner lang,[10] kan være mættet eller umættet, og kan hæftes på funktionelle grupper indeholdende oxygen, halogener, nitrogen og svovl. Hvis en fedtsyre indeholder en dobbeltbinding er der en mulighed for enten en cis eller trans geometrisk isomerisme, som i væsentlig grad påvirker molekylets konfiguration. Cis-dobbeltbindinger får fedtsyrekæden til at bøje, hvilket styrkes yderligere med dobbeltbindinger i kæden. Tre dobbeltbindinger i 18-carbon linolensyre, den mest udbredte fedt-acyl-kæde i planters thylakoidmembraner, gør disse membraner stærkt flydende på trods af miljøets lave temperaturer,[11] og får ligeledes linolensyre til at give dominerende høje spidser i højopløsning 13-C NMR-spektre af kloroplast. Dette spiller en vigtig rolle i cellemembraners struktur og funktion.[12] De fleste naturligt forekommende fedtsyrer har en cis-konfiguration, selvom trans-formen eksisterer i nogle naturlige og delvist hydrogenerede fedt og olier.[13]
Blandt eksempler på biologisk vigtige fedtsyrer er eikosanoider, der primært stammer fra arakidonsyre og icosapentaensyre, som inkluderer prostaglandiner, leukotriener og tromboxaner. Docosahexaensyre er også vigtigt i biologiske systemer, særligt hvad angår syn.[14][15] Blandt andre større lipid-klasser i kategorien af fedtsyrer er fedtestere og fedtamider. Fedtestere omfatter vigtige biokemiske mellemprodukter såsom voksestere, fedtsyre- thioester koenzym A-derivater, fedtsyre-thioester ACP-derivater og fedtsyre-carnitiner. Fedtamiderne omfatter N-acylethanolaminer, såsom cannabinoide-neurotransmitteren anandamid.[16]
Glycerolipider
[redigér | rediger kildetekst]Glycerolipider består af mono-, di- og tri-substituerede glyceroler,[17] hvoraf de bedst kendte er fedtsyre-triestere af glycerol, kaldt triglycerider. Ordet "triacylglycerol" bruges somme tider synonymt med "triglycerid". I disse forbindelser er hver af de tre hydroxyl-grupper af glycerol esterificerede, typisk af forskellige fedtsyrer. Da de fungerer som en form for energilagring, udgør disse lipider størstedelen af fedtlageret i animalsk væv. Hydrolysen af ester-bindinger af triglycerider, samt frigørelsen af glycerol og fedtsyrer fra fedtvæv er de begyndende skridt ved metabolisering af fedt.[18]
Yderligere underklasser af glycerolipider repræsenteres ved glycosylglyceroler, som er karakteriseret ved tilstedeværelsen af en eller flere sukkerrremanenser vedhæftet glycerol via en glykosidbinding. Blandt eksempler på strukturer i denne kategori er digalactosyldiacylglyceroler, der findes i plantemembraner,[19] og seminolipider fra pattedyrs sædceller.[20]
Glycerofosfolipider
[redigér | rediger kildetekst]Glycerofosfolipider, normalt omtalt som fosfolipider, er overalt i naturen, og er nøglekomponenter i cellers lipiddobbeltlag,[21] såvel som medvirkende ved metabolisme og cellesignalering.[22] Nervevæv (heriblandt hjernen) indeholder relativt store mængder glycerofosfolipider, og forandringer i deres sammensætning er blevet implicerede ved diverse neurologiske sygdomme.[23] Glycerofosfolipider kan underinddeles i særskilte klasser baseret på af glycerolrygradens polære hovedgruppes sn-3-position i eukaryoter og eubakterier, eller sn-1-position hos arkæer.[24]

Blandt eksempler på glycerofosfolipider, der findes i biologisk membran er fosfatidylcholin (også kendt som PC, GPCho eller lecitin), fosfatidyletanolamin (PE eller GPEtn) og fosfatidylserin (PS eller GPSer). Udover at tjene som en primær komponent i cellemembran og bindingssteder for intra- og intercellulære proteiner, er nogle glycerofosfolipider i eukaryotiske celle, såsom fosfatidylinositoler og fosfatidinsyrer, enten forløbere for, eller selv, membran-deriverede second messengers.[25] Typisk er en eller begge disse hydroxylgrupper aculerede med lang-kædede fedtsyrer, men der findes også alkyl-forbundne og 1Z-alkenyl-forbundne (plasmalogen) glycerofosfolipider, såvel som dialkylæter-varianter arkæer.[26]
Sphingolipider
[redigér | rediger kildetekst]Sphingolipider er en kompliceret familie af forbindelser[27] som har en fælles strukturel egenskab, en sphingoidbase-rygrad, der syntetiseres de novo fra aminosyren serin og en lang-kædet fedt-acyl-CoA, og konverteres derefter til ceramider, fosfosphingolipider, glycosphingolipider og andre forbindelser. Pattedyrs store sphingoidbase omtales normalt som sphingosin. Ceramider (N-acyl-sphingoidbaser) er en stor underklasse af sphingoidbase-derivativer med en amid-forbundet fedtsyre. Fedtsyrerne er typisk mættede eller enkeltumættede med en kædelængde på mellem 16 og 26 carbonatomer.[28]

Pattedyrs store fosfosphingolipider er sphingomyeliner (ceramidfosfocholiner),[29] hvorimod insekter hovedsageligt indeholder ceramidfosfoætanolaminer[30] og svampe har fytoceramid-fosfoinositoler og mannose-indeholdende hovedgrupper.[31] Glycosphingolipider er en forskelligartet molekylefamilie bestående af en eller flere sukkerremanenser forbundet via en glykosidbinding til sphingoidbasen. Blandt eksempler på disse er de simple og komplekse glykosphingolipider såsom cerebrosider og gangliosider.
Sterollipider
[redigér | rediger kildetekst]Sterollipider, såsom kolesterol og dets derivativer, er en vigtig bestanddel i membranlipider,[32] sammen med glycerofosfolipiderne og sphingomyelinerne. Steroider, der alle afledes af den samme kondenserede fire-ringede kernestruktur , har forskellige biologiske roller som hormoner og signalerende molekyler. Blandt steroiderne med 18 carbon (C18) er østrogen-familien, hvorimod C19-steroiderne omfatter androgenerne såsom testosteron og androsteron. C21-underklassen omfatter progestogenerne såvel som glukokortikoiderne og mineralokortikoiderne.[33] Secosteroiderne, bestående af forskellige former for vitamin D, er karakteriseret ved spaltning af B-ringen i kernestrukturen.[34] Blandt andre eksempler på steroler er galdesyrer og deres konjugater,[35] som i pattedyr er oxiderede kolesterol-derivater og syntetiseres i leveren. Plante-ækvivalenterne er fytosteroler, såsom β-sitosterol, stigmasterol og brassicasterol; hvoraf den sidstnævnte forbindelse også bruges som en biomarkør for algevækst.[36] Den dominerende sterol i svampe-cellemembraner er ergosterol.[37]
Prenollipider
[redigér | rediger kildetekst]Prenollipider syntetiseres fra fem-carbon-enhedforløberne isopentenylpyrofosfat og dimetylallylpyrofosfat, der centralt produceres via mevalonsyre (MVA)-veje.[38] De simple isoprenoider (lineære alkoholer, difosfater, etc.) dannes af successive tilsætning af C5-enheder, og klassificeres i henhold til et antal af disse terpenenheder. Strukturer, der indeholder mere end 40 carboner kendes som polyterpener. Karotenoider er vigtige simple isoprenoider, der fungerer som antioxidanter og forløbere for vitamin A.[39] En anden biologisk vigtig molekyleklasse eksemplificeres af quinoner og hydroquinoner, som indeholder en isoprenoid-hale vedhæftet en quinonoid-kerne af ikke-isoprenoid oprindelse.[40] Vitamin E og vitamin K, såvel som ubiquinoner, er eksempler fra denne klasse. Prokaryoter syntetiserer polyprenoler (kaldet bactoprenoler) hvorved den sidste isoprenoid-enhed vedhæftet oxygen forbliver umættet, hvorimod den sidste isoprenoid reduceres i animalske polyprenoler (dolicholer).[41]
Sakkarolipider
[redigér | rediger kildetekst]
Sakkarolipider er forbindelser, hvori fedtsyrer forbindes direkte til en sukker-rygrad, og danner strukturer der er kompatible med membran-dobbeltlag. I sakkarolipiderne erstatter en monosakkarid glycerol-rygraden i glycerolipider og glycerofosfolipider. De mest udbredte sakkarolipider er acylerede glukosamin-forløbere til Lipid A-komponenten af lipopolysakkarider i Gram-negative bakterier. Almindelige lipid A-molekyler er disakkarider af glukosamin, som er derivatiserede med op til syv fedt-acyl-kæder. Den minimale lipopolysakkarid der kræves for vækst i E. coli er Kdo2-Lipid A, en hexa-acyleret disakkarid af glukosamin, som er glykosyleret med to 3-deoxy-D-manno-octulosonsyre (Kdo)-remanens.[42]
Polyketider
[redigér | rediger kildetekst]Polyketider syntetiseres af polymerisering af acetyl og propionyl-underenheder af klassiske enzymer, såvel som iterative og multimodulære enzymer, der deler samme mekanistiske egenskaber som fedtsyresyntaser. De består af et stort antal sekundære metabolitter og naturprodukter fra dyr, planter, bakterier, svampe og maritime kilder, og har en stor strukturel forskelligartethed.[43][44] Mange polyketider er cykliske mokeyler, hvis rygrad ofte er yderligere modificeret af glycosylering, metylering, hydroxylering, oxidering og/eller andre processer. Mange ofte anvendte anti-mikrobielle, anti-parasitiske eller cancerbekæmpende antibiotika er polyketider eller derivater heraf, såsom erytromyciner, tetracykliner, avermectiner og tumorbekæmpende epothiloner.[45]
Biologiske funktioner
[redigér | rediger kildetekst]Membraner
[redigér | rediger kildetekst]Eukaryotiske celler er inddelte i membran-bundne organeller, der udfører forskellige biologiske funktioner. Glycerofosfolipider er den centralt strukturelle komponent i biologisk membran, såsom den cellulære plasmamembran og organellernes intracellulære membraner. Mens det især er glycerofosfolipider der er den store komponent i biologiske membraner, kan man også finde andre ikke-glyceride lipidkomponenter såsom sphingomyelin og steroler (centralt kolesterol i animalsk cellemembran).[46] I planter og alger er galactosyldiacylglyceroler,[47] og sulfoquinovosyldiacylglycerol,[19] som mangler en fosfatgruppe, vigtige membrankomponenter i kloroplast og relaterede organeller, og er de mest udbredte lipider i fotosyntetisk væv, blandt andet i højere planter, alger og visse bakterier.
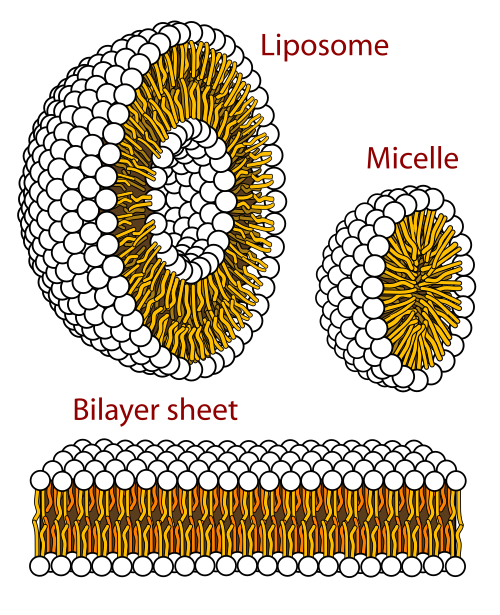
En biologisk membran er en form for lamellar-fase lipiddobbeltlag. Dannelsen af lipiddobbeltlag er en energetisk foretrukket proces når glycerofosfolipiderne er i et vandigt miljø.[48] Dette kendes som den hydrofobiske effekt. I et vandigt system stiller lipidernes polære hoveder sig mod det polære, vandige miljø, mens de hydrofobiske haler minimerer deres kontakt med vand, sætter sig sammen i klynger og danner en vesikel; ved lipidens kritiske koncentration kan denne biofysiske interaktion resultere i dannelsen af miceller, liposomer eller lipiddobbeltlag. Andre aggregeringer kan også observeres og danner en del af den amfifile (lipid)-opførsels polymorfisme. Faseopførsel er et område, der studeres indenfor biofysik.[49][50] Miceller og dobbeltlag dannes i det polære medie ved en proces kendte som den hydrofobiske effekt.[51] Når et lipofilt eller amfifilt stof opløses i et polært miljø bliver de polære molekyler (dvs. vand i en vandig opløsning) mere ordnet omkring det opløste kipofile stof, siden de polære molekyler ikke kan danne hydrogenbindinger til det amfifile stofs lipofile områder. Så i et vandigt miljø vil vandmolekylerne danne et ordnet klatrat-bur rundt omkring den opløste lipofile molekyle.[52]
Omdannelsen af lipider til protocelle-membraner er et vigtigt skridt i modeller for abiogenese, livets oprindelse.[53]
Energilagring
[redigér | rediger kildetekst]Triglycerider, opbevaret i fedtvæv, er en vigtig form for energilagring i både dyr og planter. Fedtcellerne er designet til uafbrudt syntese og nedbrydning af triglycerider i dyr, med nedbrydningen centralt kontrolleret af aktiveringen af det hormon-sensitive enzym lipase.[54] Den fuldstændige oxidering af fedtsyrer leverer et højt kalorieindhold, omkring 9 kcal/g, sammenlignet med 4 kcal/g for nedbrydningen af kulhydrater og proteiner. Trækfugle, der skal flyve lange afstande uden føde, bruger lagret energi fra triglycerider som brændstof til deres flyvning.[55]
Signalering
[redigér | rediger kildetekst]I de seneste år er der fremkommet beviser der indikerer at lipidsignaling er en central del af cellesignaling.[56][57] Lipidsignalering kan ske via aktivering af G-protein-koblede eller kernereceptorer, og medlemmer af flere forskellige lipidkategorier er blevet identificeret som signaleringsmolekyler og cellulære budbringere.[58] Blandt disse er sphingosin-1-fosfat, en ceramid-deriveret sphingolipid, der er en potent budbringermolekyle, der er involveret i regulering af calciummobilisering,[59] cellevækst og apoptose;[60] diacylglycerol (DAG) og fosfatidylinositol-fosfater (PIP) involveret i calcium-medieret aktivering af proteinkinase C;[61] prostaglandiner, som er en type fedtsyrer deriveret fra eicosanoid involveret ved betændelse og immunitet;[62] steroidehormoner såsom østrogen, testosteron og kortisol, som modulerer en række funktioner, såsom reproduktion, metabolisme og blodtryk; og oxysteroler såsom 25-hydroxy-kolesterol som er lever-X-receptor agonister.[63] Fosfatidylserinlipider vides at være involveret ved signalering til fagocytose af apoptotiske celler og/eller cellestykker. De gør dette ved at blive eksponeret for cellemembranens ekstracellulære front efter inaktiveringen af flippaser, som placerer dem eksklusivt på den cytosoliske side og aktiveringen af scramblaser, som "scrambler" fosfolipidernes orientering. Efter dette kan andre celler genkende fosfatidylserinerne og fagocytoserer cellerne eller cellefragmenterne.[64]
Yderligere læsning
[redigér | rediger kildetekst]- Smør, margarine eller planteolier? Fedt er en kompliceret sag. Videnskab.dk
- Bhagavan NV (2002). Medical Biochemistry. San Diego: Harcourt/Academic Press. ISBN 978-0-12-095440-7.
- Devlin TM (1997). Textbook of Biochemistry: With Clinical Correlations (4th udgave). Chichester: John Wiley & Sons. ISBN 978-0-471-17053-2.
- Stryer L, Berg JM, Tymoczko JL (2007). Biochemistry (6th udgave). San Francisco: W.H. Freeman. ISBN 978-0-7167-8724-2.
{{cite book}}: CS1-vedligeholdelse: Flere navne: authors list (link) - van Holde KE, Mathews CK (1996). Biochemistry (2nd udgave). Menlo Park, California: Benjamin/Cummings Pub. Co. ISBN 978-0-8053-3931-4.
Fodnoter
[redigér | rediger kildetekst]- ^ Maitland, Jr Jones (1998). Organic Chemistry. W W Norton & Co Inc (Np). s. 139. ISBN 978-0-393-97378-5.
- ^ Stryer et al., p. 328.
- ^ a b c Stryer et al., p. 330.
- ^ a b c Fahy E, Subramaniam S, Murphy RC, Nishijima M, Raetz CR, Shimizu T, Spener F, van Meer G, Wakelam MJ, Dennis EA (2009). "Update of the LIPID MAPS comprehensive classification system for lipids". Journal of Lipid Research. 50 (S1): S9-14. doi:10.1194/jlr.R800095-JLR200. ISSN 0022-2275. PMC 2674711. PMID 19098281.
- ^ Subramaniam S, Fahy E, Gupta S, Sud M, Byrnes RW, Cotter D, Dinasarapu AR, Maurya MR (2011). "Bioinformatics and systems biology of the lipidome". Chemical Reviews. 111 (10): 6452-6490. doi:10.1021/cr200295k. PMC 3383319. PMID 21939287.
- ^ Mashaghi S, Jadidi T, Koenderink G, Mashaghi A (2013). "Lipid nanotechnology". International Journal of Molecular Sciences. 14 (2): 4242-4282. doi:10.3390/ijms14024242. PMC 3588097. PMID 23429269.
- ^ Michelle A, Hopkins J, McLaughlin CW, Johnson S, Warner MQ, LaHart D, Wright JD (1993). Human Biology and Health. Englewood Cliffs, New Jersey, USA: Prentice Hall. ISBN 978-0-13-981176-0.
- ^ Vance JE, Vance DE (2002). Biochemistry of Lipids, Lipoproteins and Membranes. Amsterdam: Elsevier. ISBN 978-0-444-51139-3.
- ^ Brown HA, red. (2007). Lipodomics and Bioactive Lipids: Mass Spectrometry Based Lipid Analysis. Methods in Enzymology. Vol. 423. Boston: Academic Press. ISBN 978-0-12-373895-0.
- ^ Hunt SM, Groff JL, Gropper SA (1995). Advanced Nutrition and Human Metabolism. Belmont, California: West Pub. Co. s. 98. ISBN 978-0-314-04467-9.
- ^ Yashroy RC. (1987). "13C NMR studies of lipid fatty acyl chains of chloroplast membranes". Indian Journal of Biochemistry and Biophysics. 24 (6): 177-178.
- ^ Devlin, pp. 193–195.
- ^ Hunter JE. (2006). "Dietary trans fatty acids: review of recent human studies and food industry responses". Lipids. 41 (11): 967-992. doi:10.1007/s11745-006-5049-y. PMID 17263298.
- ^ Furse, Samuel (2011-12-02). "A Long Lipid, a Long Name: Docosahexaenoic Acid". The Lipid Chronicles.
- ^ "DHA for Optimal Brain and Visual Functioning". DHA/EPA Omega-3 Institute.
- ^ Fezza F, De Simone C, Amadio D, Maccarrone M (2008). "Fatty acid amide hydrolase: a gate-keeper of the endocannabinoid system". Subcellular Biochemistry. Subcellular Biochemistry. 49: 101-132. doi:10.1007/978-1-4020-8831-5_4. ISBN 978-1-4020-8830-8. PMID 18751909.
- ^ Coleman RA, Lee DP (2004). "Enzymes of triglyceride synthesis and their regulation". Progress in Lipid Research. 43 (2): 134-176. doi:10.1016/S0163-7827(03)00051-1. PMID 14654091.
- ^ van Holde and Mathews, pp. 630–31.
- ^ a b Hölzl G, Dörmann P (2007). "Structure and function of glycoglycerolipids in plants and bacteria". Progress in Lipid Research. 46 (5): 225-243. doi:10.1016/j.plipres.2007.05.001. PMID 17599463.
- ^ Honke K, Zhang Y, Cheng X, Kotani N, Taniguchi N (2004). "Biological roles of sulfoglycolipids and pathophysiology of their deficiency". Glycoconjugate Journal. 21 (1-2): 59-62. doi:10.1023/B:GLYC.0000043749.06556.3d. PMID 15467400.
- ^ "The Structure of a Membrane". The Lipid Chronicles. Hentet 2011-12-31.
- ^ Berridge MJ, Irvine RF (1989). "Inositol phosphates and cell signalling". Nature. 341 (6239): 197-205. doi:10.1038/341197a0. PMID 2550825.
- ^ Farooqui, A. A.; Horrocks, L. A.; Farooqui, T (2000). "Glycerophospholipids in brain: Their metabolism, incorporation into membranes, functions, and involvement in neurological disorders". Chemistry and physics of lipids. 106 (1): 1-29. doi:10.1016/S0009-3084(00)00128-6. PMID 10878232.
- ^ Ivanova PT, Milne SB, Byrne MO, Xiang Y, Brown HA (2007). "Glycerophospholipid identification and quantitation by electrospray ionization mass spectrometry". Methods in Enzymology. Methods in Enzymology. 432: 21-57. doi:10.1016/S0076-6879(07)32002-8. ISBN 978-0-12-373895-0. PMID 17954212.
- ^ van Holde and Mathews, p. 844.
- ^ Paltauf F. (1994). "Ether lipids in biomembranes". Chemistry and Physics of Lipids. 74 (2): 101-139. doi:10.1016/0009-3084(94)90054-X. PMID 7859340.
- ^ Merrill AH, Sandhoff K. (2002). "Sphingolipids: metabolism and cell signaling", Ch. 14 in New Comprehensive Biochemistry: Biochemistry of Lipids, Lipoproteins, and Membranes, Vance, D.E. and Vance, J.E., eds. Elsevier Science, NY, ISBN 978-0-12-182212-5.
- ^ Devlin, pp. 421–422.
- ^ Hori T, Sugita M (1993). "Sphingolipids in lower animals". Progress in Lipid Research. 32 (1): 25-45. doi:10.1016/0163-7827(93)90003-F. PMID 8415797.
- ^ Wiegandt H. (1992). "Insect glycolipids". Biochimica et Biophysica Acta. 1123 (2): 117-126. doi:10.1016/0005-2760(92)90101-Z. PMID 1739742.
- ^ Guan X, Wenk MR (2008). "Biochemistry of inositol lipids". Frontiers in Bioscience. 13 (13): 3239-3251. doi:10.2741/2923. PMID 18508430.
- ^ Bach D, Wachtel E (2003). "Phospholipid/cholesterol model membranes: formation of cholesterol crystallites". Biochim Biophys Acta. 1610 (2): 187-197. doi:10.1016/S0005-2736(03)00017-8. PMID 12648773.
- ^ Stryer et al., p. 749.
- ^ Bouillon R, Verstuyf A, Mathieu C, Van Cromphaut S, Masuyama R, Dehaes P, Carmeliet G (2006). "Vitamin D resistance". Best Practice & Research Clinical Endocrinology & Metabolism. 20 (4): 627-645. doi:10.1016/j.beem.2006.09.008. PMID 17161336.
- ^ Russell DW. (2003). "The enzymes, regulation, and genetics of bile acid synthesis". Annual Review of Biochemistry. 72: 137-174. doi:10.1146/annurev.biochem.72.121801.161712. PMID 12543708.
- ^ Villinski JC, Hayes JM, Brassell SC, Riggert VL, Dunbar R (2008). "Sedimentary sterols as biogeochemical indicators in the Southern Ocean". Organic Geochemistry. 39 (5): 567-588. doi:10.1016/j.orggeochem.2008.01.009.
- ^ Deacon J. (2005). Fungal Biology. Cambridge, Massachusetts: Blackwell Publishers. s. 342. ISBN 978-1-4051-3066-0.
- ^ Kuzuyama T, Seto H (2003). "Diversity of the biosynthesis of the isoprene units". Natural Product Reports. 20 (2): 171-183. doi:10.1039/b109860h. PMID 12735695.
- ^ Rao AV, Rao LG (2007). "Carotenoids and human health". Pharmacological Research: the Official Journal of the Italian Pharmacological Society. 55 (3): 207-216. doi:10.1016/j.phrs.2007.01.012. PMID 17349800.
- ^ Brunmark A, Cadenas E (1989). "Redox and addition chemistry of quinoid compounds and its biological implications". Free Radical Biology & Medicine. 7 (4): 435-477. doi:10.1016/0891-5849(89)90126-3. PMID 2691341.
- ^ Swiezewska E, Danikiewicz W (2005). "Polyisoprenoids: structure, biosynthesis and function". Progress in Lipid Research. 44 (4): 235-258. doi:10.1016/j.plipres.2005.05.002. PMID 16019076.
- ^ a b Raetz CR1, Garrett TA, Reynolds CM, Shaw WA, Moore JD, Smith DC Jr; et al. (2006). "Kdo2-Lipid A of Escherichia coli, a defined endotoxin that activates macrophages via TLR-4". The Journal of Lipid Research. 47 (5): 1097-1111. doi:10.1194/jlr.M600027-JLR200. PMID 16479018.
{{cite journal}}: CS1-vedligeholdelse: Flere navne: authors list (link) - ^ Walsh CT. (2004). "Polyketide and nonribosomal peptide antibiotics: modularity and versatility". Science. 303 (5665): 1805-1810. doi:10.1126/science.1094318. PMID 15031493.
- ^ Caffrey P, Aparicio JF, Malpartida F, Zotchev SB (2008). "Biosynthetic engineering of polyene macrolides towards generation of improved antifungal and antiparasitic agents". Current Topics in Medicinal Chemistry. 8 (8): 639-640. doi:10.2174/156802608784221479. PMID 18473889.
- ^ Minto RE, Blacklock BJ (2008). "Biosynthesis and function of polyacetylenes and allied natural products". Progress in Lipid Research. 47 (4): 233-306. doi:10.1016/j.plipres.2008.02.002. PMC 2515280. PMID 18387369.
- ^ Stryer et al., pp. 329–331.
- ^ Heinz E. (1996). "Plant glycolipids: structure, isolation and analysis", pp. 211–332 in Advances in Lipid Methodology, Vol. 3. W.W. Christie (ed.). Oily Press, Dundee. ISBN 978-0-9514171-6-4
- ^ Stryer et al., pp. 333–334.
- ^ van Meer G, Voelker DR, Feigenson GW (2008). "Membrane lipids: where they are and how they behave". Nature Reviews Molecular Cell Biology. 9 (2): 112-124. doi:10.1038/nrm2330. PMC 2642958. PMID 18216768.
- ^ Feigenson GW. (2006). "Phase behavior of lipid mixtures". Nature Chemical Biology. 2 (11): 560-563. doi:10.1038/nchembio1106-560. PMC 2685072. PMID 17051225.
- ^ Wiggins PM. (1990). "Role of water in some biological processes". Microbiological Reviews. 54 (4): 432-449. PMC 372788. PMID 2087221.
- ^ Raschke TM, Levitt M (2005). "Nonpolar solutes enhance water structure within hydration shells while reducing interactions between them". Proceedings of the National Academy of Sciences of the United States of America. 102 (19): 6777-6782. doi:10.1073/pnas.0500225102. PMC 1100774. PMID 15867152.
- ^ Segré D, Ben-Eli D, Deamer D, Lancet D (2001). "The Lipid World" (PDF). Origins of Life and Evolution of Biospheres. 31 (1-2): 119-145. doi:10.1023/A:1006746807104. PMID 11296516. Arkiveret fra originalen (PDF) 11. september 2008. Hentet 14. juni 2016.
- ^ Brasaemle DL. (2007). "Thematic review series: adipocyte biology. The perilipin family of structural lipid droplet proteins: stabilization of lipid droplets and control of lipolysis". Journal of Lipid Research. 48 (12): 2547-2559. doi:10.1194/jlr.R700014-JLR200. PMID 17878492.
- ^ Stryer et al., p. 619.
- ^ Wang X. (2004). "Lipid signaling". Current Opinion in Plant Biology. 7 (3): 329-236. doi:10.1016/j.pbi.2004.03.012. PMID 15134755.
- ^ Dinasarapu AR, Saunders B, Ozerlat I, Azam K, Subramaniam S (2011). "Signaling gateway molecule pages—a data model perspective". Bioinformatics. 27 (12): 1736-1738. doi:10.1093/bioinformatics/btr190. PMC 3106186. PMID 21505029.
- ^ Eyster KM. (2007). "The membrane and lipids as integral participants in signal transduction". Advances in Physiology Education. 31 (1): 5-16. doi:10.1152/advan.00088.2006. PMID 17327576.
- ^ Hinkovska-Galcheva V, VanWay SM, Shanley TP, Kunkel RG (2008). "The role of sphingosine-1-phosphate and ceramide-1-phosphate in calcium homeostasis". Current Opinion in Investigational Drugs. 9 (11): 1191-1205. PMID 18951299.
- ^ Saddoughi SA, Song P, Ogretmen B (2008). "Roles of bioactive sphingolipids in cancer biology and therapeutics". Subcellular Biochemistry. Subcellular Biochemistry. 49: 413-440. doi:10.1007/978-1-4020-8831-5_16. ISBN 978-1-4020-8830-8. PMC 2636716. PMID 18751921.
- ^ Klein C, Malviya AN (2008). "Mechanism of nuclear calcium signaling by inositol 1,4,5-trisphosphate produced in the nucleus, nuclear located protein kinase C and cyclic AMP-dependent protein kinase". Frontiers in Bioscience. 13 (13): 1206-1226. doi:10.2741/2756. PMID 17981624.
- ^ Boyce JA. (2008). "Eicosanoids in asthma, allergic inflammation, and host defense". Current Molecular Medicine. 8 (5): 335-349. doi:10.2174/156652408785160989. PMID 18691060.
- ^ Bełtowski J. (2008). "Liver X receptors (LXR) as therapeutic targets in dyslipidemia". Cardiovascular Therapy. 26 (4): 297-316. doi:10.1111/j.1755-5922.2008.00062.x. PMID 19035881.
- ^ Biermann M, Maueröder C, Brauner JM, Chaurio R, Janko C, Herrmann M, Muñoz LE (2013). "Surface code—biophysical signals for apoptotic cell clearance". Physical Biology. 10 (6): 065007. doi:10.1088/1478-3975/10/6/065007. PMID 24305041.
Eksterne links
[redigér | rediger kildetekst]| Wikimedia Commons har medier relateret til: |